the Creative Commons Attribution 4.0 License.
the Creative Commons Attribution 4.0 License.
 | 08 Dec 2025
| 08 Dec 2025
Taxonomy, systematics, and valve ultrastructure of the Rhopalodiales (Bacillariophyta: Bacillariophyceae) taxa from Swan Lake, Oregon, and a checklist of the group in the USA
John P. Kociolek
Megan Greenwood
Aimee Caye G. Chang
Sarah E. Hamsher
Scott R. Miller
Jingchun Li
We describe species of Rhopalodiales found in samples from Swan Lake, Oregon, deposited in the Tempère and Peragallo Collection in the Museum National d'Histoire Naturelle (Paris) and the Academy of Natural Sciences (Philadelphia). A total of eight different taxa have been recognized, and these are detailed with light and, in some cases, scanning electron microscopy. Six of the species are of the genus Epithemia sensu stricto. Within this genus, we describe one species new to science and make two new combinations. Species found in the Swan Lake material belong to the Turgida (one species), Argus (one species), Sorex (two species), and Adnata (two species) groups of Epithemia. Variation is described relative to the course and position of the raphe branches, the structure and organization of the areolae, and the central area. Two species of Rhopalodia sensu stricto are reported, including one newly described species. A checklist of the Epithemia and Rhopalodia species reported from the USA is also presented.
- Article
(20507 KB) - Full-text XML
- BibTeX
- EndNote




Members of the Rhopalodiales are known to, in some instances, dominate the periphyton community in Cladophora and higher plants (Power et al., 2009; DeYoe et al., 1992; Kelly, 2003; Stancheva et al., 2013). While they may be numerically dominant in systems with low N : P ratios (either very oligotrophic systems where the bedrock may be comprised of high phosphorus volcanic rock or in systems where phosphorus has been anthropogenically or experimentally introduced; e.g., Fairchild et al., 1985), the number of different taxa comprising these communities is relatively modest. Collections from fossil localities in France reveal the largest diversity of species from this order. For example, Heribaud (1920) reported 21 different Epithemia taxa and 20 different Rhopalodia taxa from travertine localities in Auvergne, France. Localities identified as “Joursac” and “Ceyssat” were reported to contain 14 and 13 taxa of Rhopalodiales (Héribaud, 1903; Tempère and Peragallo, 1908, 1909), and Tempère and Peragallo (1909) report 12 taxa from Loch Kinnard, Scotland.
In the USA, the total number of Epithemia and Rhopalodia taxa reported is 70. However, accounting for homotypic synonyms (16), the total number is 54 (see Table 1). Of these, 15 were described to be from the United States originally. Tempère and Peragallo (1911) reported 13 taxa from a fossil locality reported as “Lost Springs Ranch”, but the locality with the largest number of reported taxa of Rhopalodiales is Swan Lake, Oregon. Tempère and Peragallo (1909) reported 14 taxa from this locality. While the location was treated by Patrick and Reimer (1975) as a recent habitat, investigations of other taxa in the genus Gomphoneis Cleve by Kociolek and Stoermer (1988) suggested that it was a fossil locality. There is an extant habitat with that name near Klamath Lake, Oregon (42°19′28′′ N, 121°36′51′′ W), but there are also several diatomites in the region (Meyers and Newcomb, 1952). Of the 14 taxa reported by Tempère and Peragallo (1909), 4 were validly described as new (as Epithemia hyndmannii var. capitata Tempère and H.Peragallo; Epithemia truncata M. Peragallo, E. trunctata var. debilis Tempère & H.Peragallo, and E. westermannii var. stricta Tempère & H.Peragallo; Servant-Vildary et al. 2001). Patrick and Reimer (1975) investigated three of the new, validly described Epithemia taxa in Tempère and Peragallo's slides (1909, edition II, slides 365, 366) from Swan Lake.
Table 1Checklist of reported Rhopalodiales taxa from the United States. L indicates lectotype; ANSP indicates Academy of Natural Sciences of Philadelphia; CAS indicates California Academy of Sciences; COLO indicates University of Colorado, Boulder: T&P(2) 366 indicates the slide from the Diatomées du Monde Entier.
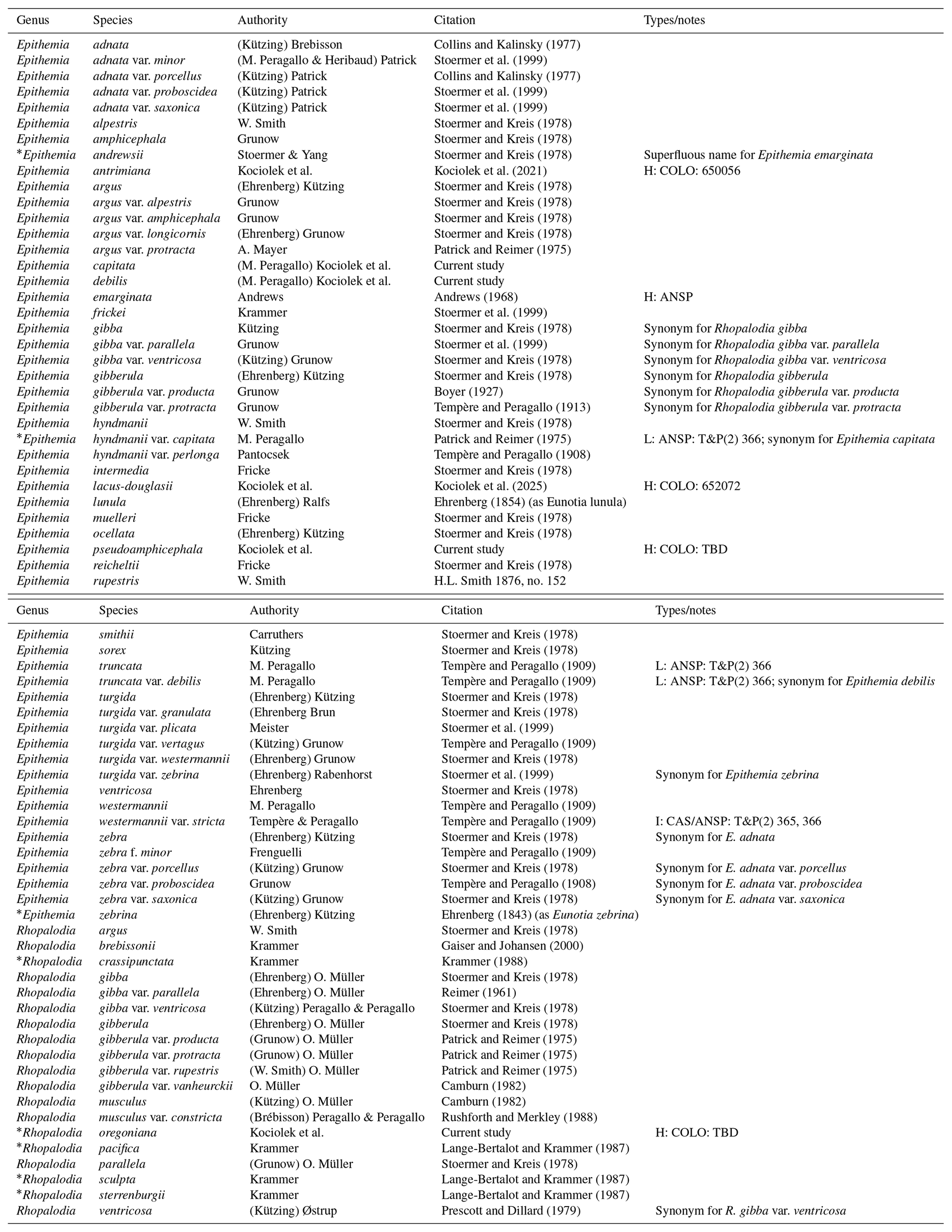
* indicates the species described from the United States.
In the context of an ongoing taxonomic revision of the Rhopalodiales (e.g., Kociolek and Van De Vijver, 2023; Kociolek et al., 2024, 2025), we review both the newly discovered and the previously reported taxa from Swan Lake. The purpose of the present report is to describe their morphology and to clarify their systematic position.
2.1 Samples
We obtained raw material from the Museum National d'Histoire Naturelle, Department of Geological Sciences, which is the original collection of Tempère and Peragallo used to make slides for their exsiccatae “Diatomées du Monde Entier” (Servant-Vildary et al., 2001).
Raw material was also obtained from The Academy of Natural Sciences, Philadelphia. The material was labeled “Swan Lake, Terry 3410”. This material was previously used by Kociolek and Stoermer (1988) in their consideration of endemic species of the genus Gomphoneis Cleve from that locality.
2.2 Processing
From these raw samples, permanent light microscope preparations were made by mixing some of the material with distilled water; air-drying that mixture onto coverslips; and, after drying, mounting the coverslips onto glass slides with Hyrax. Light microscopy (LM) observations were made with an Olympus BX-51 microscope using differential interference contrast (DIC) optics. For scanning electron microscopy (SEM), coverslips with dried material were attached to an aluminum stub with carbon tape. Stubs were sputter-coated with 4 nm of platinum using a Cressington 108 sputter coater and were examined using a Hitachi SU3500 VPSEM (Hitachi High-Tech America, Inc., Schaumburg, Illinois) at an accelerating voltage of 15 kV at the COSINC facility at the University of Colorado, Boulder.
We follow the terminology used by Sims (1983) and Kociolek et al. (2025) for Epithemia species. For Rhopalodia species, the terminology of Krammer (1988a, b) is followed.
Holotypes are deposited at the J.P. Kociolek Collection at the University of Colorado, Boulder (COLO). Isotypes are deposited at the Academy of Natural Sciences, Philadelphia, and the Natural History Museum, London.
-
Genus Epithemia Brébisson ex Kützing
-
Epithemia capitata (Tempère & M. Peragallo) Kociolek et al., comb. nov., stat. nov.
Plate 1, figs. 1–13; Plate 2, figs. 1–16.
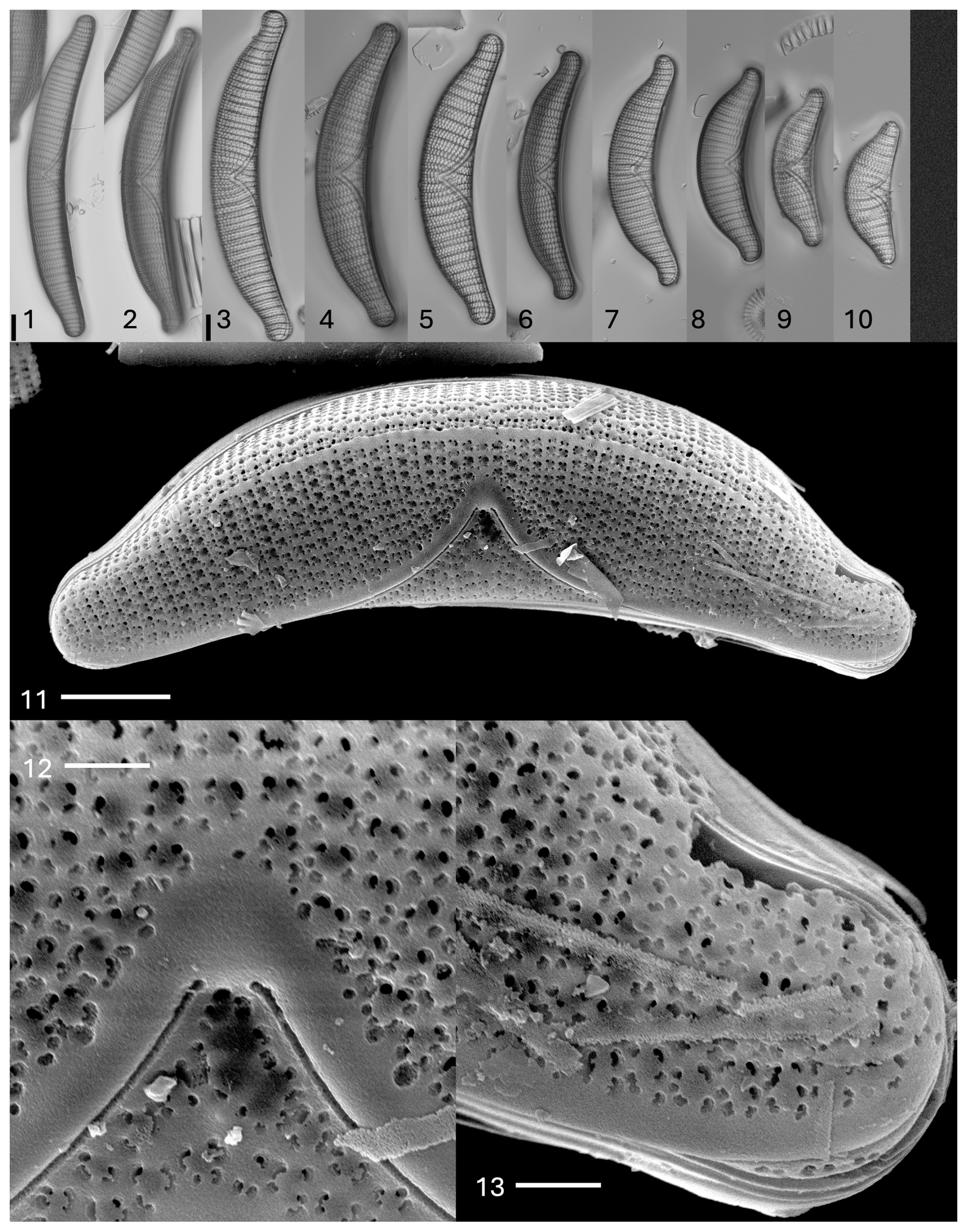
Plate 1Epithemia capitata (Tempère & M. Peragallo) Kociolek et al., comb. nov., stat. nov. Light microscopy, valve views. Size diminution series. The scale bar shown in (1) is 10 µm for (1) and (2); the scale bar shown in (3) is 10 µm for (3)–(10). (11–13) SEM. External views. (11) Valve view. Dorsal ridge, proximal raphe ends, and complex areolae are evident. The scale bar is 10 µm. (12) Center of valve face. The scale bar is 2 µm. (13) Apex of the valve. Raphe is positioned on the ventral margin and mantle. The scale bar is 2 µm.

Plate 2Epithemia capitata (Tempère & M. Peragallo) Kociolek et al., comb. nov., stat. nov. (14–16) SEM. Internal views. (14) Valve view. Costate fibulae are evident. A large central area is present. The canal raphe is narrow. The scale bar is 10 µm. (15) The central area is comprised of a cave-like area. The scale bar is 2 µm. (16) Areolar openings and costate fibulae are evident. The scale bar is 2 µm.
Basionym. Epithemia hyndmanii var. capitata Tempère & Peragallo 1909, p. 194, slide nos. 365, 366 (non 1910 as indicated in Patrick & Reimer 1975)
Description. The frustules are planar. The valves are asymmetrical in relation to the apical axis. The valves show a strongly convex dorsal margin and a concave ventral margin, with apices that are capitate and slightly recurved dorsally, with lengths of 68–180 µm and breadths of 14–24 µm. The canal raphe is positioned at the center of each apex; it curves towards the ventral margin and then towards the valve face, extending about one-half of the way between the ventral and dorsal margins. Raphe branches are evident on the valve face. The portules are small and indistinct, about the size of the areolae. The costate fibulae are narrow and radiate and number four to six per 10 µm. The areolae are rectangular and radiate, with two striae between adjacent fibulae, numbering 7–11 per 10 µm. The septa are at the poles only.
In the SEM, there is a ridge of silica extending along most of the dorsal margin. Areolae on the valve face appear as irregular openings, but on the dorsal mantle, there are domes over the areolae that are interconnected (Plate 1, figs. 11–13). The proximal raphe ends are within hyaline areas, and the branches are bordered by a thin ridge of silica (Plate 1, fig. 12). The valve exterior has areolae with four external openings for each opening internally (Plate 1, figs. 12 and 13). The raphe is present on the valve face at the apices, runs along the ventral margin, and then extends back onto the valve face about one-half of the way across the length of the valve. The raphe ends extend about two-thirds of the way from the ventral margin towards the dorsal margin (Plate 1, figs. 11–13).
Internally, the costate fibulae are prominent, while secondary costae are rare (Plate 2, fig. 14). There are between two to four rows of areolae between adjacent costate fibula (Plate 2, fig. 15). The raphe canal has very small portulae. The central nodule is demarcated by shortened but prominent costate fibulae (Plate 2, fig. 15). Girdle bands have extensions onto the fibulae. Each areolar opening internally produces four external openings (Plate 2, fig. 16).
Comments. Patrick's description (in Patrick and Reimer, 1975, p. 186) is mostly accurate, except for the size range of this species and the placement of the central nodule. Peragallo's original description (in Tempère and Peragallo, 1909, p. 194) was succinct: “Peu arque et a extremities recurvees et capitate” [Slightly arched and with curved, capitate end].
As a note, the name Epithemia hyndmanii (hyndmannii) var. truncata M. Peragallo in Tempère & Peragallo was listed by both Mills (1934, p. 657) and Van Landingham (1969, p. 1452) as being from Tempère and Peragallo slides 365 and 366 (Swan Lake). The name was never proposed by Tempère and Peragallo, and it must have been a mistake introduced by Mills and continued by Van Landingham. It is a nomen nudum.
-
Epithemia debilis (Tempère & M. Peragallo) Kociolek et al. comb. nov., stat. nov.
Plate 3, figs. 17–36; Plate 4, figs. 37–40
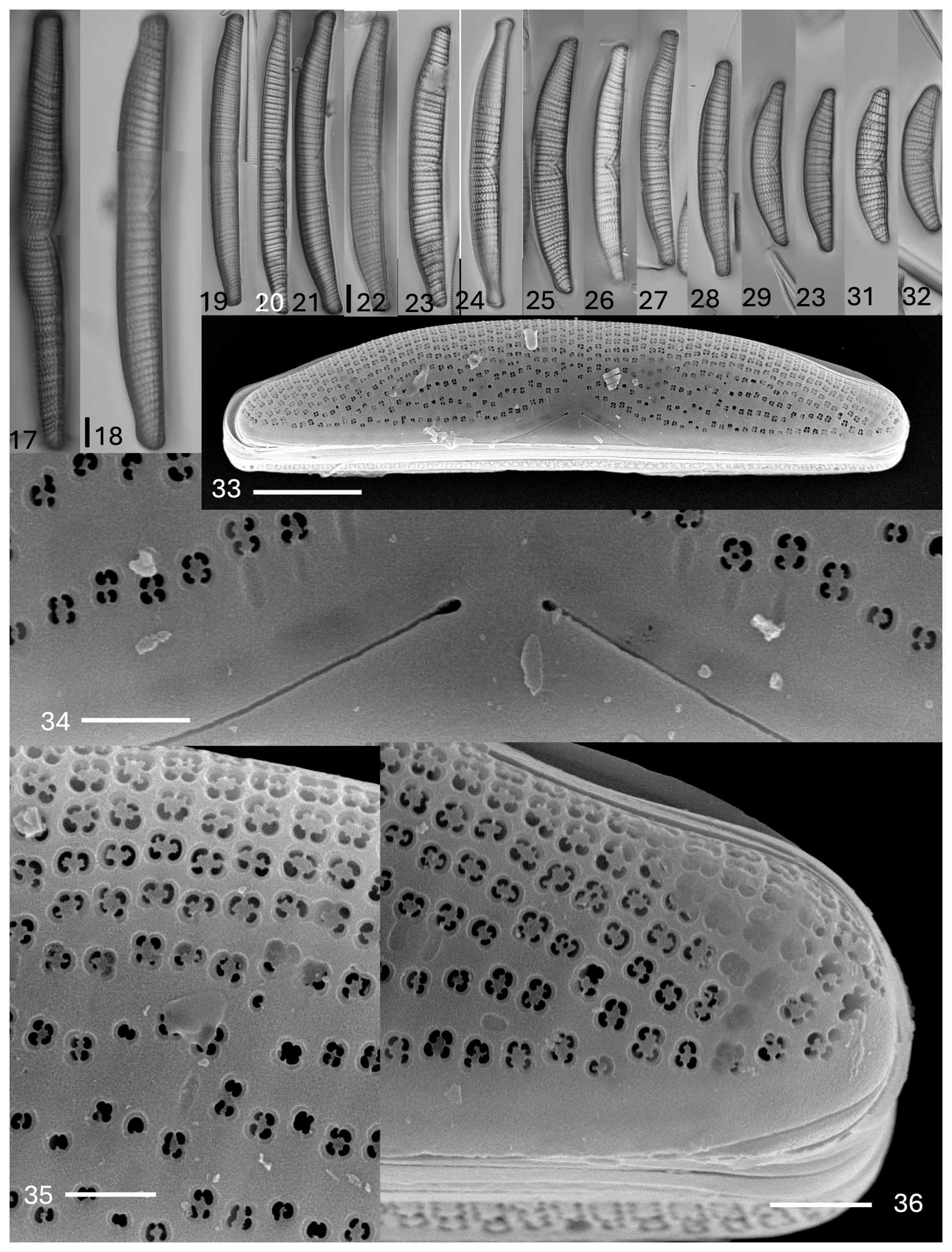
Plate 3Epithemia debilis (Tempère & M. Peragallo) Kociolek et al., comb. nov., stat. nov. (17–32) Light microscopy, valve views. Size diminution series. Figure (17) is of an initial valve. The scale bar shown in (17) is 10 µm for (17) and (18); the scale bar shown in (22) is 10 µm for (19)–(32). (33–36) SEM. External views. (33) Entire valve view. Striae have denser areolae towards the dorsal margin as compared to the ventral margin. Areolae are absent ventrally in the proximal raphe ends. The scale bar is 10 µm. (34) Central area showing proximal raphe ends. Areolae are sunken with four openings. The scale bar is 2 µm. (35) Dorsal margin showing the density of areolae in striae. The scale bar is 2 µm. (36) Apex, showing the density of areolae. The scale bar is 2 µm.
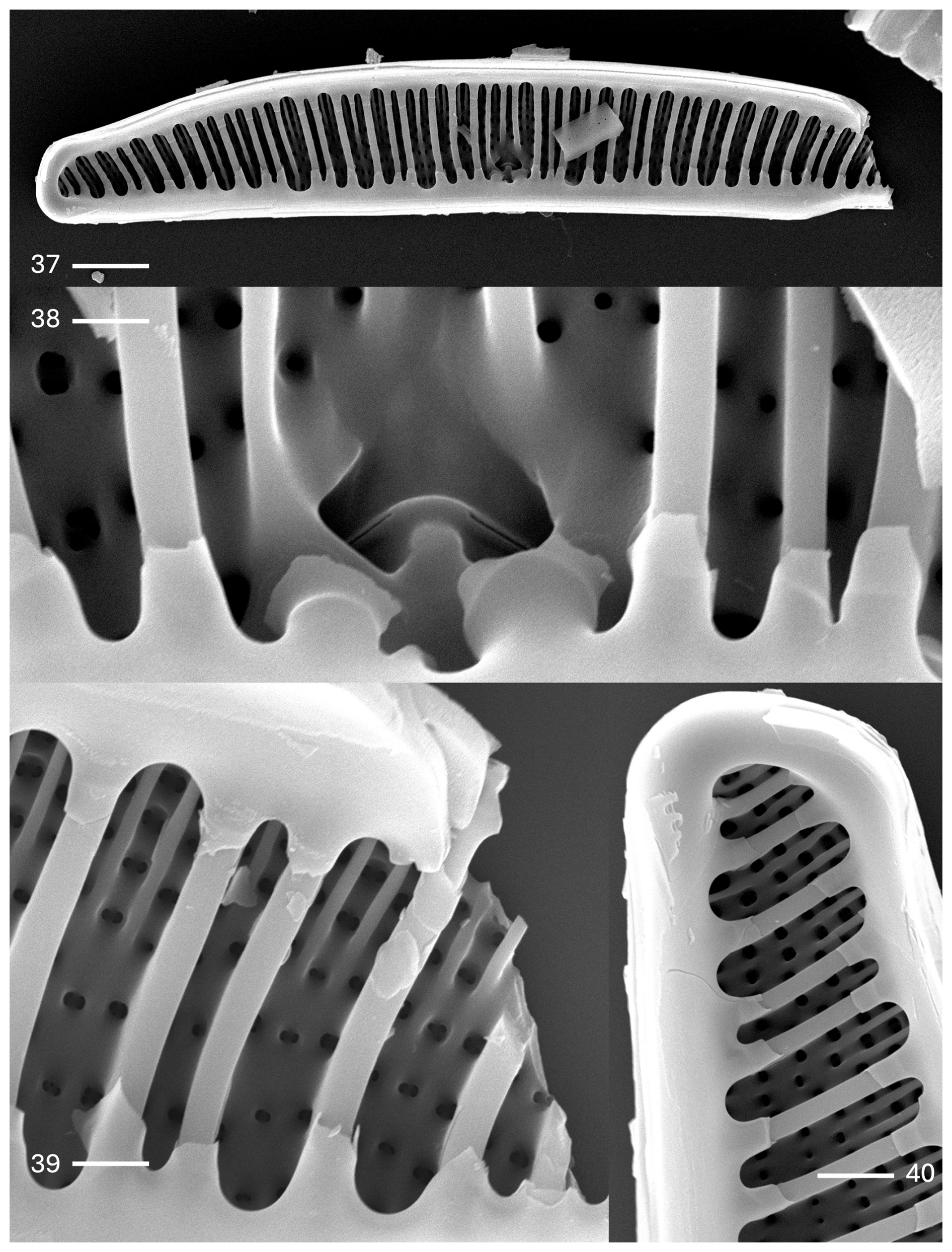
Plate 4Epithemia debilis (Tempère & M. Peragallo) Kociolek et al., comb. nov., stat. nov. (37–40) SEM. Internal views. (37) Nearly the entire valve. Girdle bands with extensions overlaying the fibulae are evident. Costate fibulae may extend entirely across the valve face or nearly so. The scale bar is 10 µm. (38) Central area. The internal proximal raphe ends are discontinuous. Arrows indicate extensions from the girdle band. The scale bar is 1 µm. (39) Broken portion of valve showing the round internal areolar openings, thick fibulae, and extensions from the girdle band. The scale bar is 1 µm. (40) Apex of the valve with a small septum evident. The scale bar is 1 µm.
Basionym. Epithemia truncata var. debilis Tempère & M. Pergallo, 1909, p. 194.
Synonym. Epithemia westermanii var. stricta M. Peragallo in Tempère & Peragallo, 1909, p. 194.
Description. The frustules are planar. The valves are asymmetrical in relation to the apical axis. The dorsal margin is convex, and the ventral margin is straight, with apices that are protracted to be slightly capitate to somewhat truncate. The lengths are 52–160 µm, and the breadths are 12–15 µm. The canal raphe extends from the middle portion of the apices, runs along the ventral margin, and extends onto the valve face near the central portion of the valve to about one-quarter of the way towards the dorsal margin. The portulae are indistinct. A raphe is evident, with external proximal ends that are dilated. The costate fibulae are radiate, numbering four to six (up to 7 near the apices) per 10 µm. There are two to three striae between adjacent fibulae, with areolae being denser near the dorsal margin and less dense towards the ventral margin. The striae number 8 to 12 per 10 µm. There is little to no ornamentation near the ventral margin below the central nodule. The areolae are round to rectangular in shape. There are septa at the poles, with extensions from the girdle bands present at the fibulae.
In the SEM, the valve face has striae comprised of areolae that become less dense towards the center of the valve and towards the ventral margin (Plate 3, figs. 33–36). The raphe is within a hyaline axial area (Plate 3, fig. 34). The raphe is on the valve face at the apices and near the center of the valve, extending about one-quarter of the way from the ventral margin towards the dorsal margin (Plate 3, figs. 33 and 34). There is no ornamentation on the ventral side of the raphe. Proximal raphe ends are slightly dilated (Plate 3, fig. 34). The areolae are in small, shallow depressions with two to four (mostly) openings evident, occluded by short volae (Plate 3, figs. 35 and 36).
Internally, the prominent costate fibulae are dense; they may not extend all the way across the valve face (Plate 4, figs. 37–40). The central nodule is prominent, positioned within an elliptical area. The raphe is discontinuous across the central nodule (Plate 4, fig. 38). The girdle bands have extensions that partially cover the costate fibulae (Plate 4, figs. 39 and 40). Septa are present at the apices (Plate 4, fig. 40).
Comments. Peragallo's description of this taxon is as follows: “plus gréle et moins franchement tronque que le type, les granules s'espacecent vers le bord central et sont moins fortement marqués” [thinner and less strongly truncated than the type, the areolae are spaced out towards the central edge and less strongly evident]. There is a much broader size diminution series present in the Swan Lake material than the 120–130 µm length given by Patrick and Reimer (1975) and in the original description by Peragallo. This taxon is an endemic to Swan Lake.
Epithemia westermannii var. stricta var. nov. is described by Tempère & Peragallo (1909, p. 194) as “De meme forme mais plus étroit que le type” [same shape but narrower than the type]; Epithemia proboscidea, Epithemia turgida, and E. turgida var. granulata, reported by Tempère and Peragallo, could not be verified as present in the Swan Lake material. They all may represent longer specimens of this species.
-
Epithemia truncata M. Peragallo in Tempère & Peragallo, 1909
Plate 5, figs. 41–60
(non 1910, as indicated in Patrick & Reimer, 1975)
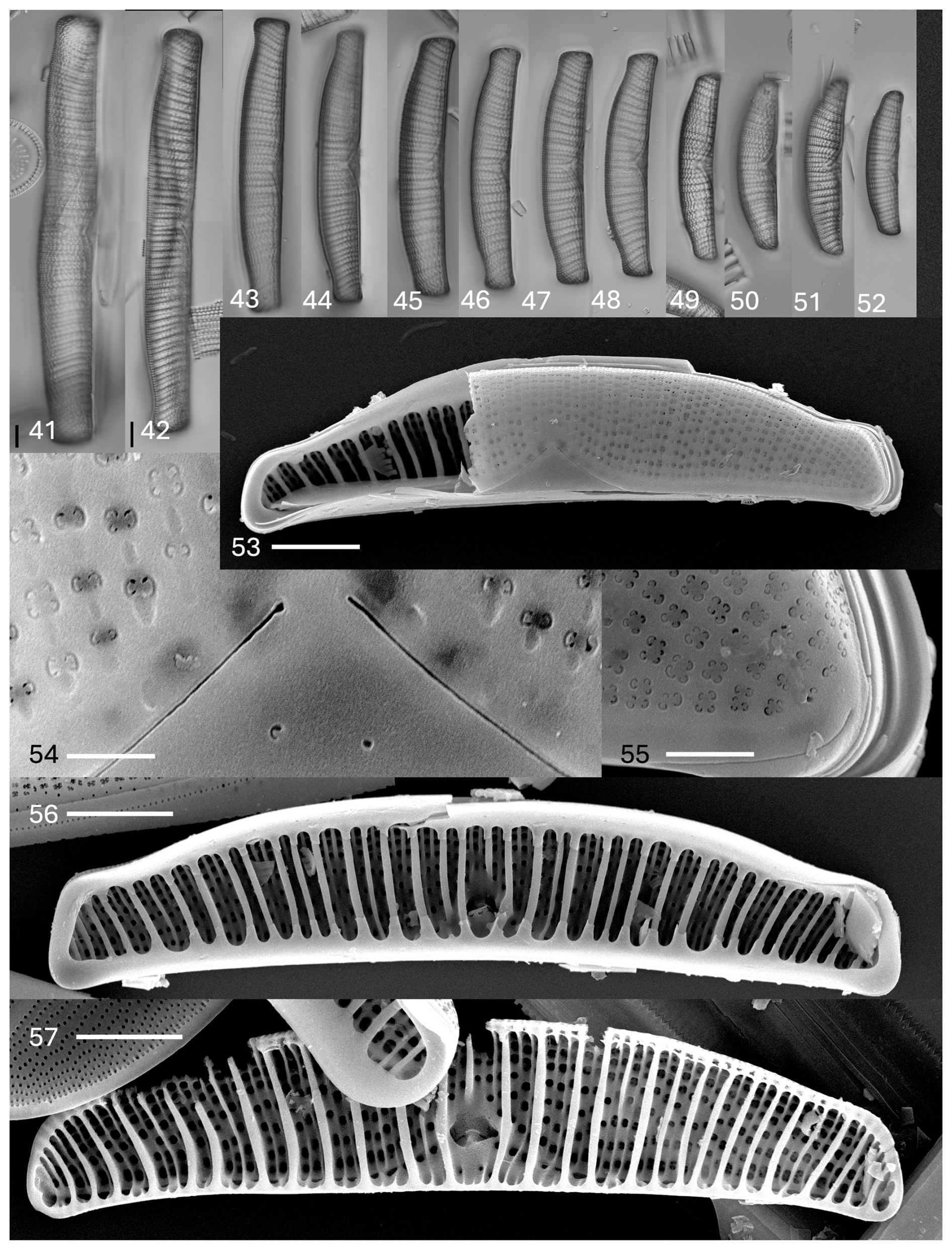
Plate 5Epithemia truncata M. Peragallo in Tempère & Peragallo. (41–52) Light microscopy, valve views. Size diminution series. Figure (41) shows an initial valve. The scale bar shown in (41) is 10 µm, and the scale bar shown in (42) is 10 µm for (42)–(52). (53–57) SEM. (53–55) External views. (53) Partial valve showing striation and proximal raphe ends. The scale bar is 10 µm. (54) Central area with proximal raphe ends evident. The areolae are nearly absent ventrally at the proximal raphe ends. The areolae may have two to four openings. The scale bar is 1 µm. (55) Apex, showing raphe on the ventral margin extending onto the mantle at the valve terminus. The scale bar is 1 µm. (56, 57) Internal views. (56) Specimen has girdle bands with extensions overlaying the costate fibulae and short septa at the poles. The scale bar is 10 µm. (57) Specimen without girdle bands. Costate fibulae and canal raphe are evident. The scale bar is 10 µm.
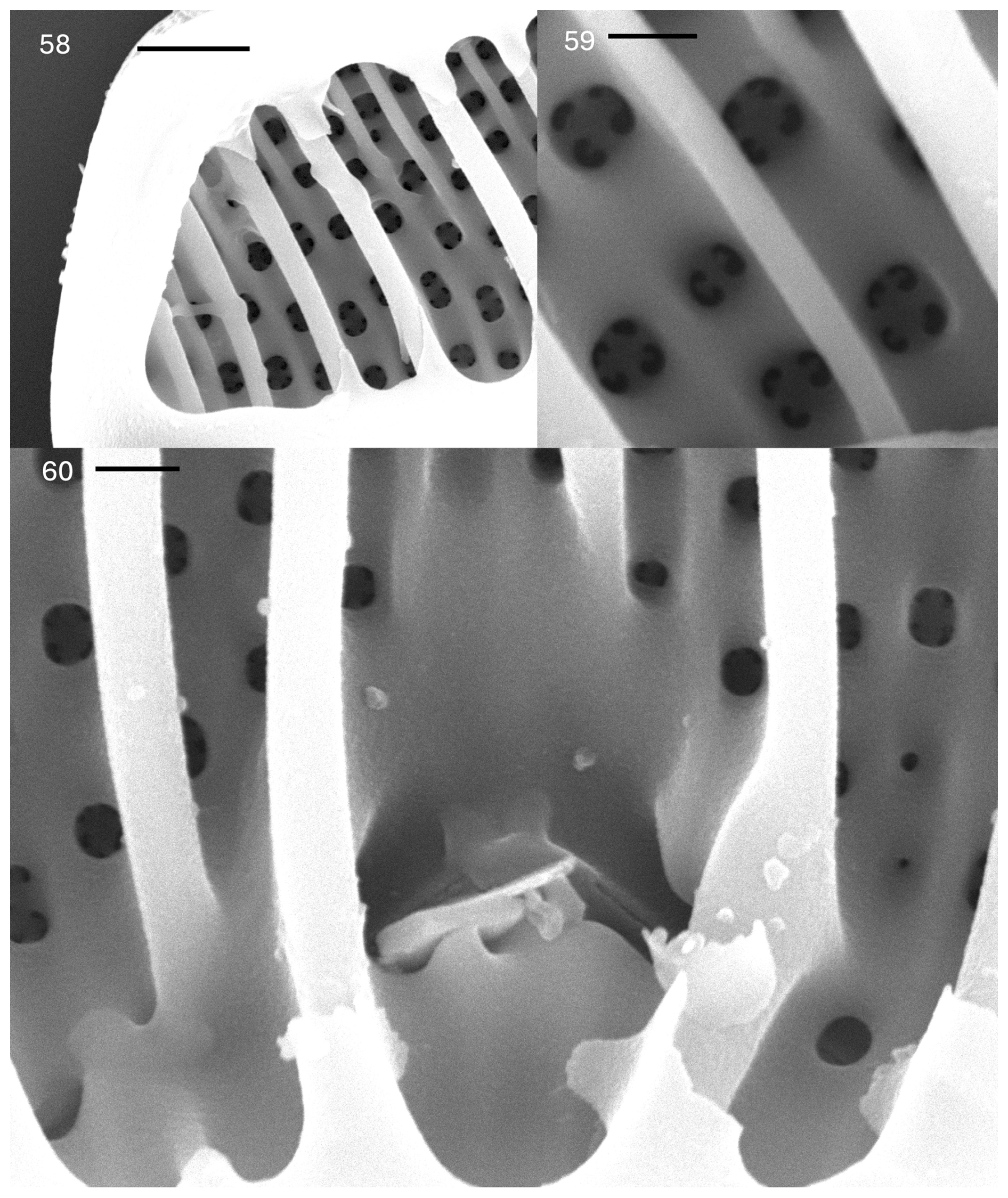
Plate 6Epithemia truncata M. Peragallo in Tempère & Peragallo. (58–60) SEM. Internal views. (58) Apex of the valve showing short septum and round internal openings of the areolae. The scale bar is 2.5 µm. (59) Valve face showing round internal areolar openings with two to four openings externally. The scale bar is 1.0 µm. (60) Central area, with proximal raphe ends that are discontinuous. The scale bar is 1.0 µm.
In the Catalogue of Diatom Names (Fourtanier & Kociolek, 2011), there is the name Epithemia hyndmanii (hyndmannii) var. truncata M. Peragallo in Tempère & Peragallo 1909, p. 194, nos. 365, 366. This name was never formally proposed by Tempère & Peragallo. The name was introduced in the Mills Index (1934, p. 657) and continued in Van Landingham's catalogue (1969, p. 1452). No description has been offered; thus, it is a nomen nudum.
Description. The frustules are planar. The valves are asymmetrical in relation to the apical axis. The dorsal margin is convex, the ventral margin is straight to very weakly concave, and the apices are separated from the body of the valve and are distinctly truncate. The lengths are 77–190 µm, and the breadths are 18–20 µm. A canal raphe is evident, extending from the apices to the ventral margin and extending onto the valve face near the center of the valve. The position on the valve face ends at about one-quarter of the way from the ventral margin to the dorsal margin. The portules are indistinct, with openings smaller than the areolae. A raphe branch is evident, with external proximal ends that are dilated. The costate fibulae are radiate, numbering three to five per 10 µm in size. There are striae with denser areolae towards the dorsal margin and striae with less dense and irregularly spaced areolae towards the ventral margin, as well as two to three striae between adjacent fibulae, numbering 8–10 per 10 µm. There is minimal ornamentation between the central nodule and the ventral margin. The areolae are squarish to round in shape. There are septa at the poles only.
In the SEM, the areolae on the valves have C-shaped openings, found in groups of two or four (rarely more) (Plate 5, figs. 53–55). The raphe extends from the ventral mantle onto the valve face only at the center, extending only about one-fifth of the distance between the ventral and dorsal margins. The external proximal raphe ends are slightly dilated (Plate 5, figs. 53 and 54). No ornamentation is present between the raphe ends and the ventral margin (Plate 5, fig. 54). The raphe can be seen at the apices on the mantle (Plate 5, fig. 55). Internally, the primary costate fibulae are evident (Plate 5, figs. 56 and 57). Girdle bands have short extensions that extend along the bases of the fibulae (Plate 5, fig. 56; Plate 6, figs. 58 and 60). The portulae of the canal raphe are small, approximately of the size of the areolar openings or smaller (Plate 6, fig. 60). The central nodule is obvious at the central opening of the canal raphe. The proximal raphe ends are evident; the raphe is discontinuous at the center (Plate 6, fig. 60). Each areolar opening expands into two to four or more openings externally. Secondary fibulae alternate with primary costate fibulae (Plate 6, fig. 59).
Comments. There is a much wider range of valve lengths in the Swan Lake material than what is indicated by either Peragallo (in Tempère and & Peragallo, 1909) or Patrick and Reimer (1975).
-
Epithemia cf. adnata
Plate 7, figs. 61–63
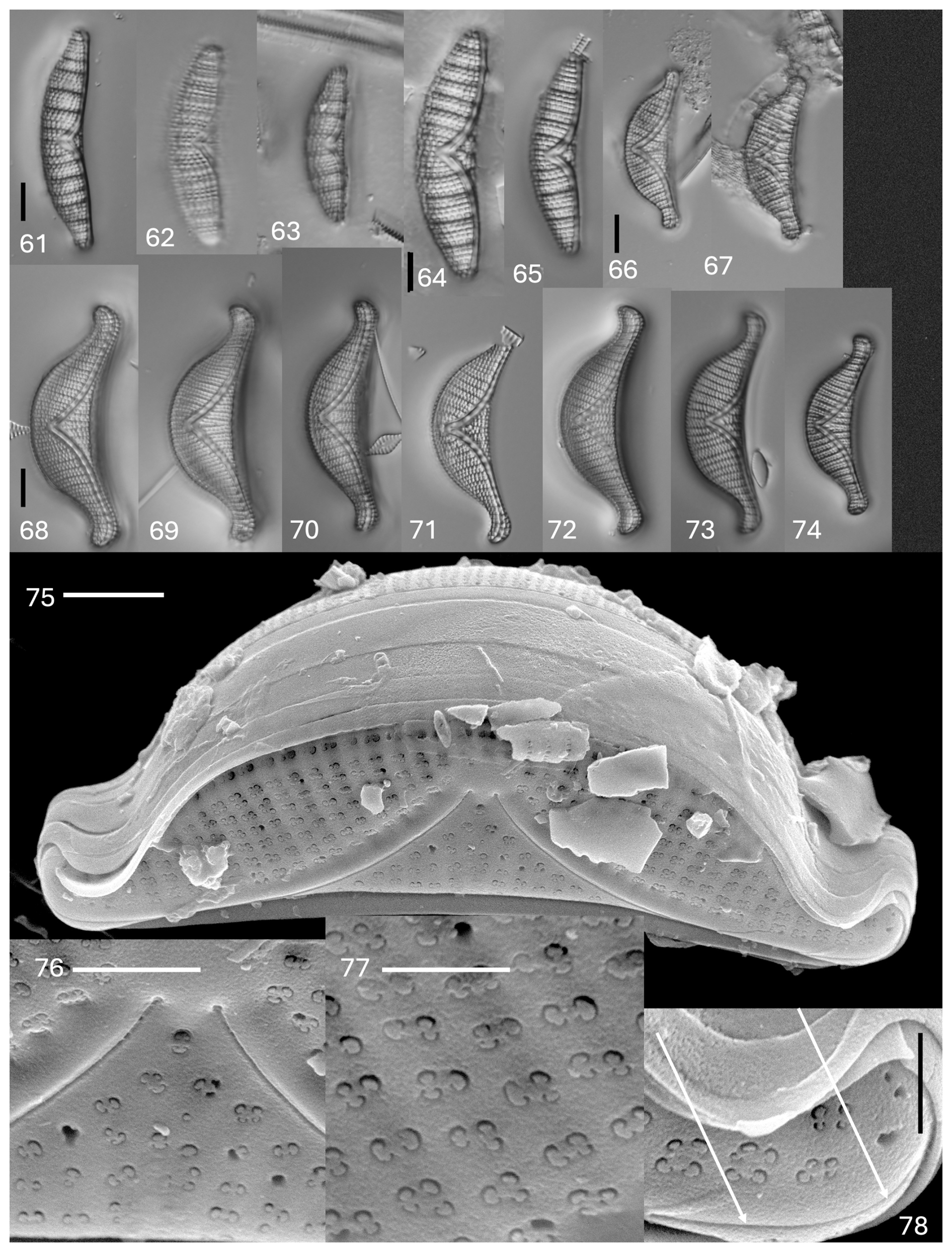
Plate 7(61–74) Light microscopy. (61–63) Epithemia cf. adnata. (64–65) Epithemia argus cf. longicornis. (66–67) Epithemia sorex Kützing. (68–74) Epithemia pseudoamphicephala Kociolek et al., sp. nov. Figure (69) shows the holotype specimen. The scale bar shown in (61) is 10 µm for (61)–(63). The scale bar shown in (64) is 10 µm for (64) and (65). The scale bar shown in (66) is 10 µm for (66) and (57). The scale bar shown in (68) is 10 µm for (68)–(74). (75–78) SEM. External views. Epithemia pseudoamphicephala Kociolek et al., sp. nov. (75) Whole valve. A biarcuate raphe is evident. The scale bar is 5 µm. (76) Central area with proximal raphe ends. The scale bar is 2 µm. (77) Valve face showing striae with areolae with two to four openings with volate occlusions. The scale bar is 1 µm. (78) Apex of the vale. Arrows indicate the raphe branch. The scale bar is 2 µm.
Description. The frustules are planar. The valves are asymmetrical in relation to the apical axis, with apices that are protracted and rounded (larger specimens) or not protracted (smaller specimens). The lengths are 29–40 µm, and the breadths are 7.0–8.5 µm. The costate fibulae are distinct, numbering three to four per 10 µm. The striae distinct, with two to five rows between adjacent fibulae, numbering 14–16 per 10 µm. There is a raphe that barely extends onto the valve face near the ventral margin at the center.
Comments. The few specimens present in the Swan Lake material approximate the type specimens of Kützing illustrated by Kociolek et al. (2024) but are slightly more slender than those present in Kützing's original material from Halle, Germany. A larger population would allow a more substantial evaluation of the status of this taxon present in Swan Lake and a better assessment of it as a new taxon (with potential description).
-
Epithemia argus cf. var. longicornis (Ehrenberg) Grunow
Plate 7, figs. 64 and 65
Description. The valves are asymmetrical in relation to the apical axis, with the dorsal margin being convex and the ventral margin being weakly concave. The apices are deflected dorsally but only slightly. The lengths are 30–37 µm, and the breadths are 7–9 µm. The raphe is highly biarcuate, extending from the valve face: there is a mantle junction up to more than halfway to the dorsal margin in the center of the valve. The raphe is distinct, and a canal and portules are evident. The costate fibulae are distinct and radiate in orientation and number three to five per 10 µm. The striae are comprised of distinct, squarish areolae, with two to four between adjacent fibulae, numbering 9–16 per 10 µm.
Comments. Specimens in the Swan Lake material are rare, as well as being smaller and more finely ornamented than what is normally reported for this taxon (Patrick and Reimer, 1975; Krammer and Lange-Bertalot, 1988). The lack of a larger population precludes us from describing these specimens as new to science.
-
Epithemia sorex Kützing
Plate 7, figs. 66–67
Description. The frustules are segment-like. The valves are distinctly asymmetrical in relation to the apical axis, with a dorsal margin that is strongly convex and a ventral margin that is weakly concave. The apices are distinctly protracted and rounded, deflected towards the ventral margin or remaining straight. The lengths are 33–45 µm, and the breadths are 9–11 µm. The raphe is strongly biarcuate, running from the apices towards the ventral margin, and then it arches strongly towards the dorsal margin, with the proximal raphe ends extending almost to the dorsal margin. The raphe canal and portules are distinct. The costate fibulae are not robust, numbering four to size per 10 µm. The striae are radiate, with two to three rows between adjacent costate fibulae. Distinctly punctate striae extend past the central area and to the ventral margin at the center. The striae number 10–15 per 10 µm. There are girdle bands with short extensions that overlap with the fibulae. Septa are present at the head pole.
Comments. The species is not common in the Terry material, though it matches well with the valve shape and size of the type specimens of Kützing (Kociolek et al., 2025).
-
Epithemia pseudoamphicephala Kociolek et al. sp. nov.
Plate 7, figs. 68–78; Plate 8, figs. 79–81
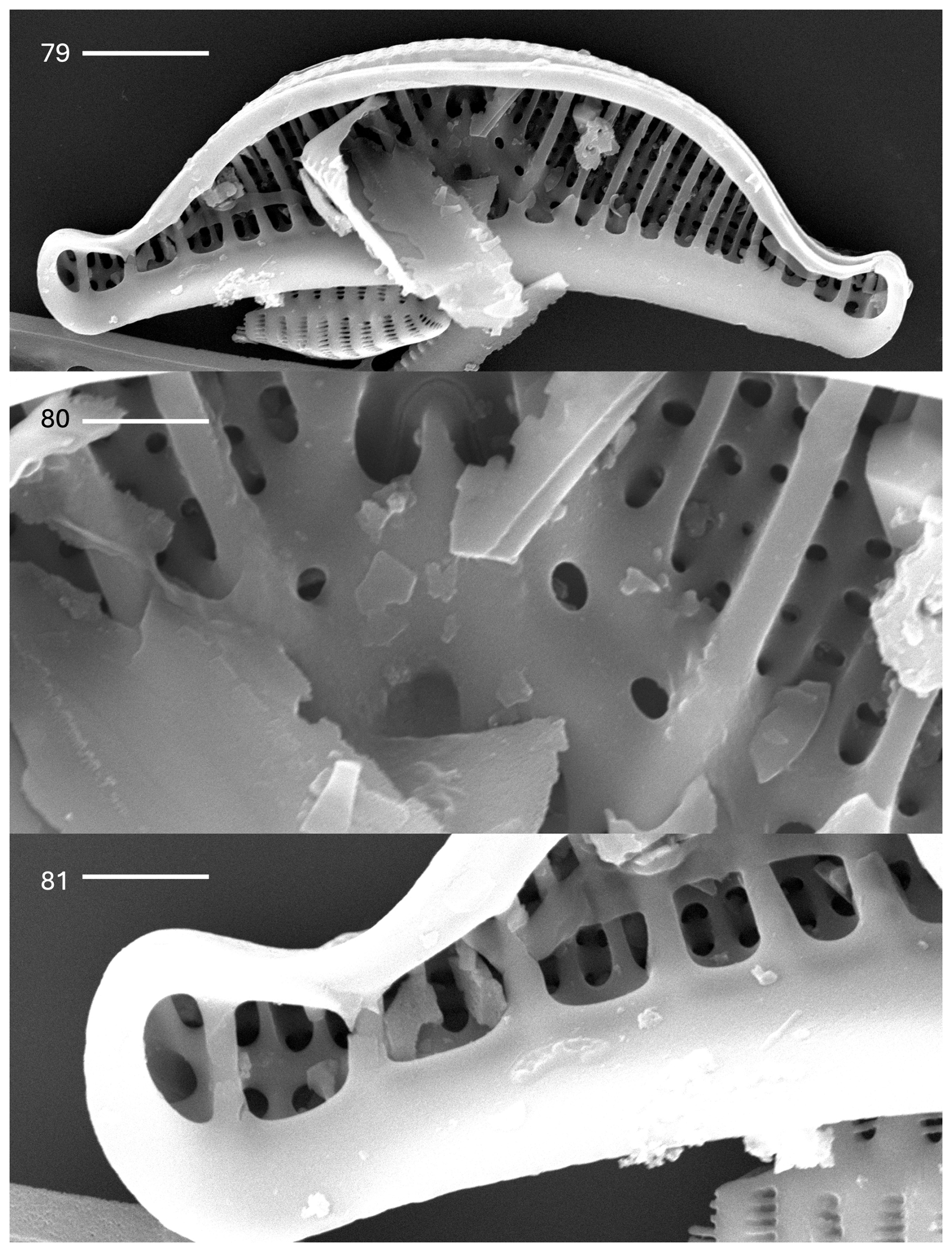
Plate 8Epithemia pseudoamphicephala Kociolek et al., sp. nov. (79–81) SEM. Internal views. (79) Whole-valve view. Extensions from girdle bands overlay costate fibulae. The raphe canal is evident. The scale bar is 5.0 µm. (80) Central area showing an inverted heart-shaped opening with a continuous raphe. The portules of the raphe canal are small. The scale bar is 2.0 µm. (81) Apex of the valve. The scale bar is 2.5 µm.
Description. The frustules are segment-like. The valves are distinctly asymmetrical in relation to the apical axis, with a dorsal margin that is strongly convex and a ventral margin that is weakly concave. The apices are distinctly protracted and rounded and are distinctly deflected towards the dorsal margin. The lengths are 40–50 µm, and the breadths are 10.5–14.0 µm. The raphe is strongly biarcuate, running from the apices towards the ventral margin, and then it arches strongly towards the dorsal margin, with the proximal raphe ends extending almost to the dorsal margin. The raphe canal and portules are distinct. The costate fibulae are not robust at four to six per 10 µm. The striae are radiate, with two to three rows between adjacent costate fibulae. Distinctly punctated striae extend past the central area and to the ventral margin at the center. The striae number 10–15 per 10 µm. There are girdle bands with short extensions that overlap with the fibulae. There are septa present at the head pole.
In the SEM, the exterior is covered by rows of areolae that are in groups of two to four (Plate 7, figs. 75–78). Each areola is occluded by a vola, giving a characteristic C shape to the opening. The valve face is smooth, without domed coverings or openings (Plate 7, figs. 75–78). The raphe is within a narrow axial area, located about mid-way on the apices, arching strongly towards the ventral margin and then arching onto the valve face and terminating in the center near the dorsal margin (Plate 7, figs. 75 and 76). Numerous girdle bands, some of the open type, are present. Internally, the raphe canal is wide and has small portules (Plate 8, figs. 79 and 80). Near the dorsal margin, the raphe can be seen to be continuous over the central nodule (Plate 8, fig. 80). Extensions from the girdle bands are nearly continuous near the apices but cover only a short segment of the costate fibulae near the center (Plate 8, fig. 81).
Holotype. Microscope slide 652076. In the JPK Collection of Diatoms at the University of Colorado, Boulder (COLO).
Isotypes. ANSP GC68121. In the Diatom Collection of the Academy of Natural Sciences (Philadelphia) and BM92464 at the Natural History Museum (London).
Comments. This species is distinguished from E. sorex by the dorsally deflected apices and slightly wider valves. Valve ultrastructural features also distinguish the two taxa. The areolae openings of both taxa are C-shaped. However, in E. pseudoamphicephala, the valve surface is flat. In extant and fossil populations of E. sorex from North and South America, respectively, the valve face has domed coverings of the areolae (Kociolek et al., 2025). In addition, E. sorex populations may have a silica ridge extending across the dorsal margin. This structure is missing in E. pseudoamphicephala.
-
Genus Rhopalodia O. Müller
-
Rhopalodia cf. gibba (Ehrenberg) O. Müller
Plate 9, figs. 82–84

Plate 9(81–91) Light microscopy. (82–84) Rhopalodia cf. gibba (Ehrenberg) O. Müller. (85–91) Rhopalodia oregoniana Kociolek et al., sp. nov. Figure (87) shows the holotype from the slide. The scale bar shown in (83) is 10 µm for (82)–(84). The scale bar shown in (85) is 10 µm for (85)–(91). (92–93) Rhopalodia oregoniana Kociolek et al., sp. nov. SEM. External views. (92) Whole valve. The raphe is evident. The scale bar is 10 µm. (93) Central area with proximal ends deflected towards the ventral margin. The C-shaped areolae are in two rows. The scale bar is 1.25 µm.
Description. The valves have a swollen center and gracefully undulate along the dorsal margin, terminating with ventrally deflected apices. The lengths are 70–110 µm, and the breadths are 10–15 µm. There are parallel costate fibulae in the middle of the valve, which are convergent at the end, at 9–11 per 10 µm. There are striae with two rows between adjacent fibulae, at 18–22 per 10 µm.
Comments. The outline of the dorsal margin is quite unlike the margin seen in some interpretations of this species, which is straighter (e.g., Patrick and Reimer, 1975; Krammer and Lange-Bertalot, 1988). A very small population of this taxon is present in the Swan Lake material. Observations on additional specimens may prove that these specimens are deserving of being recognized as new.
-
Rhopalodia oregoniana Kociolek et al. sp. nov.
Plate 9, figs. 85–95
Description. The frustules are segment-like, with numerous girdle bands. The lengths are 48.0–92.0 µm, and the breadths are 9.5–12.0 µm. There are valves with raphe canals evident along the curved (“dorsal”) edge. Portules are not evident. Margins with the raphe undulate near the center; there is no central “notch” evident. The apices are curved towards the ventral margin. The ventral margin is straight, except at the apices. There are parallel costate fibulae at the center, and there are radiate costate fibulae at the ends of the valve, at 9–11 per 10 µm. Two to three rows of striae occur between adjacent fibulae, comprised of small openings, at 13–16 per 10 µm.
In the SEM, the valve exterior is comprised of the raphe within a hyaline axial area along the dorsal margin (Plate 9, figs. 92 and 93). There is no defined keel. The proximal raphe ends are deflected towards the ventral margin (Plate 9, figs. 92 and 93), and the same is true for the distal raphe ends (Plate 10, fig. 94). In some specimens, the areolae appear to be round in shape (Plate 10, fig. 94), but, in better-preserved specimens, the areolae have volae that impart the characteristic C-shaped openings (Plate 9, fig. 93). In girdle view, the girdle bands are wide and bear a single row of poroids (Plate 10, fig. 95).
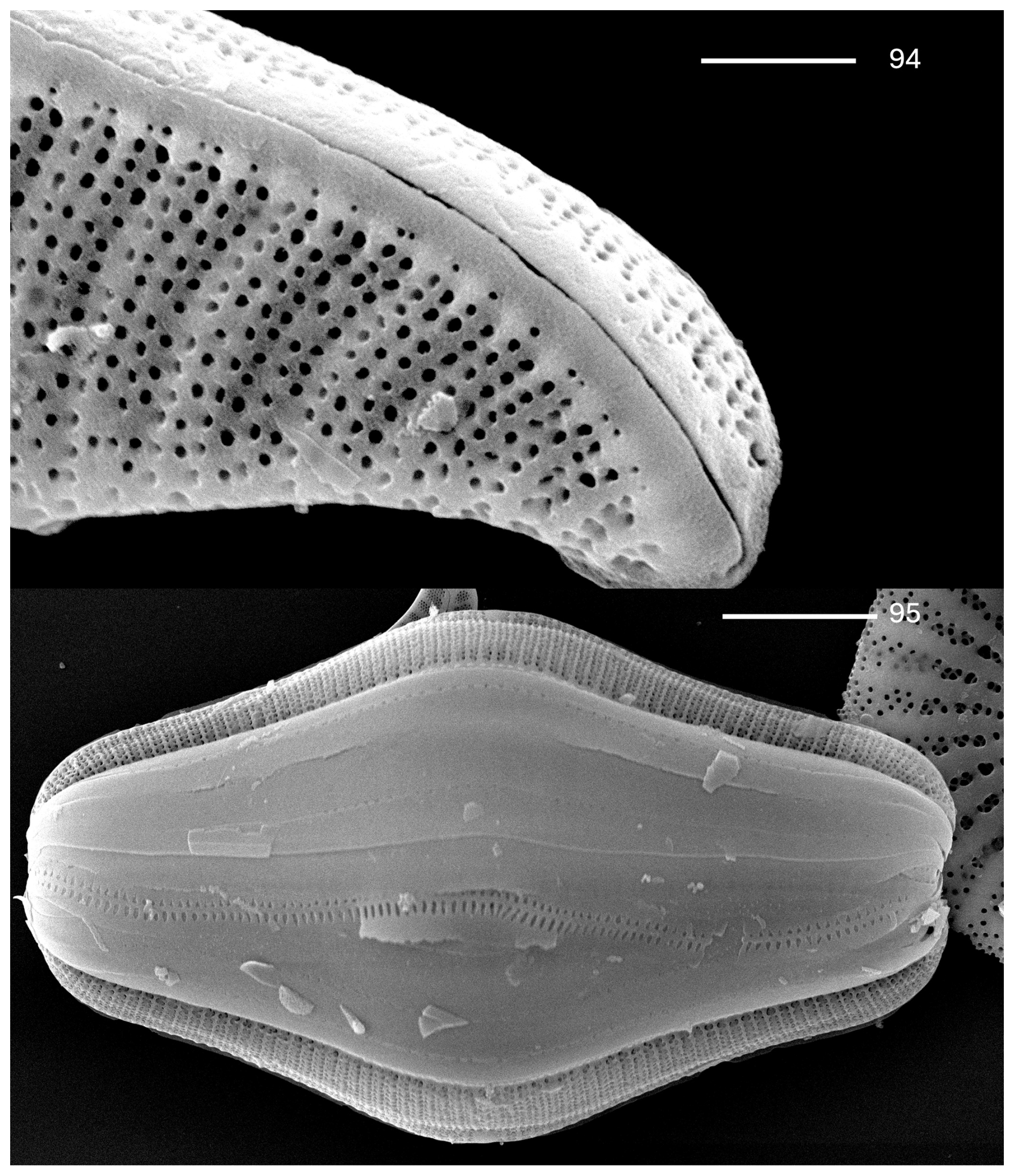
Plate 10Rhopalodia oregoniana Kociolek et al., sp. nov. (94–95) SEM. External views. (94) Apex of the valve. The distal raphe end is deflected ventrally. The raphe is within a very weakly developed keel. The scale bar is 2.5 µm. (95) Frustule, with many girdle bands evident. The scale bar is 10 µm.
Holotype. Microscope slide 652076. In the JPK Collection of Diatoms at the University of Colorado, Boulder (COLO).
Isotypes. ANSP GC68121. In the Diatom Collection of the Academy of Natural Sciences (Philadelphia) and BM92464 at the Natural History Museum (London).
Comments. This species differs from R. gibba in terms of the overall shape of the valves. We believe that these specimens were referred to as R. gibba var. ventricosa by Tempère and Peragallo (1909) in their taxon register for the Swan Lake sample. Kociolek et al. (2024) detailed the type specimens of Kützing's Epithemia ventricosa, which differs in terms of the shape of the valve and is not known from such large or small cells as seen in R. oregoniana. Krammer and Lange-Bertalot (1988) suggested that R. gibba var. ventricosa (Kützing) H. & M. Pergallo is but part of the size diminution series of the nominate variety.
We can confirm the presence of eight taxa of the Rhopalodiales from the Swan Lake material of Terry in the Collection of the Academy of Natural Sciences and in the Tempère and Peragallo Diatomees du Monde Entier, slides 365 and 366, as documented above. Six of these were previously reported by Tempère and Peragallo (1909), and two of them are new to science.
In addition, four others, namely E. westermannii var. stricta, E. proboscidea, E. turgida, and E. turgida var. granulata, could not be identified in the material from ANSP (Academy of Natural Sciences of Philadelphia) and MNHN (Muséum national d’histoire naturelle, Paris). Here, three of them are considered to be either synonyms for E. debilis or confused with it, being found among the larger individuals of the latter taxon. The record of E. turgida was likely to be a confusion with some of the smaller specimens of E. capitata. Further work on these taxa, especially the type specimens on which they are based, is warranted to determine if this suggestion of synonymy is accurate and what, if any, nomenclatural implications there may be if supported.
Five of the eight taxa found from Swan Lake appear to be endemic to this site, with these being E. pseudoamphicephala, E. truncata, E. capitata, E. debilis, and R. oregoniana. However, the number of fossil diatom sites across the western United States is incredibly large (see Eardley-Wilmot, 1928; Skinner et al., 1944; Hanna, 1956; Feth, 1964; Krebs and Bradbury, 1995; Krebs et al., 1987; Wallace et al., 2006), and there is an abundance of sites in southern Oregon alone (e.g., Smith, 1932; Moore, 1937; Newcomb, 1958; Peterson and McIntire, 1970 – this included an LM of an unknown species of Epithemia in it (see their Fig. 27-b); https://ngmdb.usgs.gov/Geolex/UnitRefs/YonnaRefs_12314.html, last access: 10 April 2025), and the diatoms of these sites have not been studied in great detail. At sites where taxonomic work has been done, most of these studies have concerned themselves with the dominant taxa. Thus, we might fully expect that the four taxa considered here to be endemic to Swan Lake are, indeed, more widely distributed.
With this work, we add two new species as additions to the Epithemia flora of the United States, making the total number of taxa reported sit at 74 and, after homotypic synonyms, leaving us with 58 taxa in total (Table 1).
Kociolek et al. (2025) proposed five distinct morphological groups within the genus Epithemia. These groups are based on the structure and course of the raphe system, the structure of the areolae, and the size and shape of the valves. Of the six Epithemia taxa from the Swan Lake locality, one species, E. capitata, is from the Turgida group. This group commonly has larger valves, a complex areolar structure, and a raphe system that is strongly biarcuate (i.e., going from the apices towards the ventral margin and then arching towards the dorsal margin, with the external proximal raphe ends reaching about half of the way to the dorsal margin). Two species are from the Sorex group, both E. sorex and the new species E. psuaeodamphicephala. In these two smaller species, the highly biarcuate raphe nearly reaches the dorsal margin. Finally, there are three species present in the Swan Lake material that are from the Adnata group. In this group, represented by E. truncata, E. debilis, and E. cf. adnata, the raphe is mostly restricted to the ventral margin. The proximal raphe ends barely extend onto the valve face near the center of the valve. There are no relatives of the Cistula or the Argus groups represented in the Swan Lake flora.
The frustular morphology of Epithemia species is complex and variable. Sims (1983) and Kobayasi and Kobayashi (1988) offered early, detailed descriptions of the valve morphology of the genus. A summary of valve ultrastructure studies on Epithemia species was offered in Kociolek et al. (2025), who also considered the valve morphology of 14 taxa. Even with the documentation of the valve ultrastructure of over 35 Epithemia taxa previously published (see Kociolek et al., 2025, Table 1), there are several morphological features seen in taxa from Swan Lake that have not previously been reported. In the endemic taxa E. debilis and E. truncata, striae have higher densities of areolae near the dorsal margin, and they become much coarser towards the ventral margin. In nearly all other Epithemia species surveyed to date, areolar density is the same throughout the length of the striae (e.g., Patrick and Reimer, 1975; Krammer and Lange-Bertalot, 1988). Also, little to no ornamentation is evident in the area ventral to the proximal raphe ends of E. debilis and E. truncata. Usually, some ornamentation is present in this region of the valves in Epithemia species (Patrick and Reimer, 1975; Krammer and Lange-Bertalot, 1988). Additionally, in E. capitata, the portulae of the canal raphe are proportionately much smaller than in other species of the genus. Portulae are usually the same size as the areolae or larger (e.g., Kociolek et al., 2025).
The two taxa reported here from Rhopalodia are part of the lineage containing R. gibba and its allies. Rhopalodia gibba is the generitype of Rhopalodia. Even at the outset of the establishment of the genus, it was recognized that Rhopalodia may harbor significant morphological diversity (Müller, 1895, 1899, 1900). Krammer (1988a, b) began to describe and recognize the morphological groups of Rhopalodia and suggested that the genus may not be a natural group of taxa. Recent studies have begun to identify unique morphologies among taxa currently assigned to the genus (e.g., Kociolek et al., 2025). Ruck et al. (2016)'s molecular study confirmed that Rhopalodia is non-monophyletic, which led them to lump members of the genus into Epithemia. Vigneshwaran et al. (2021) argued that this approach would be less taxonomically conservative than recognizing the distinct lineages of taxa assigned to Rhopalodia and would mask the morphological, ecological, and evolutionary diversity of the group. Additional studies are needed to determine if morphological data support the groupings suggested by Ruck et al. (2016), separating lineages from the generitype (R. gibba) and its allies of the genus.
This review of the Rhopalodiales from Swan Lake expands our knowledge of both taxonomic and morphological diversity within this diatom lineage. These findings support the idea that further research on the group, in fossil and recent habitats, is warranted.
All data are available in the paper.
JPK developed the conceptualization of the project. JPK and MG did the visualization of the paper and worked with SEH on the formal analysis. All of the co-authors contributed to the writing and editing of the original and revised versions of the paper.
The contact author has declared that none of the authors has any competing interests.
Publisher's note: Copernicus Publications remains neutral with regard to jurisdictional claims made in the text, published maps, institutional affiliations, or any other geographical representation in this paper. While Copernicus Publications makes every effort to include appropriate place names, the final responsibility lies with the authors. Views expressed in the text are those of the authors and do not necessarily reflect the views of the publisher.
This work was supported, in part, by grant no. 2222944 from the US National Science Foundation.
This paper was edited by Taniel Danelian and reviewed by Paula Noble and one anonymous referee.
Andrews, G. W.: New names for late Pleistocene diatom species, Journal of Paleontology, 42, 244, 1968.
Boyer, C. S.: Synopsis of North American Diatomaceae. Suppement 2. Naviculatae, Surirellatae, Proc. Acad. Natl. Sci. Phila., 79, 229–583, 1927
Camburn, K. E.: Subaerial diatom communities in eastern Kentucky, Trans. Amer, Microsc. Soc., 101, 375–87, 1982.
Collins, G. B. and Kalinsky, R. G.: Studies on Ohio diatoms, Bull. Ohio Biol. Surv., N.S., 5, 1–76, 1977.
DeYoe, H. R., Lowe, R. L., and Marks, J. C.: Effects of nitrogen and phosphorus on the endosymbiont load of Rhopalodia gibba and Epithemia turgida (Bacillariophyceae), J. Phycol., 28, 773–777, 1992.
Eardley-Wilmot, V. L.: Diatomite, its occurrence, preparation and uses, Cana. Dept. Mines, Bull., 691, 182 pp., 1928.
Ehrenberg, C. G.: Verbreitung und Einflufs des mikroskopischen Lebens in Süd-und Nord-Amerika, Abh. Königl. Akad. Wissens. Berlin, 1841, 291–445, 1843.
Ehrenberg, C. G.: Mikrogeologie. Das Erden und Felsen schaffende Wirken des unsichtbar kleinen selbstständigen Lebens auf der Erde, Leopold Voss, Leipzig, xxviii + 374 pp., 1854.
Fairchild, G. W., Lowe, R. L., and Richardson, W. B.: Algal periphyton growth on nutrient-diffusing substrates: an in situ bioassay, Ecology, 66, 465–472, 1985.
Feth, J. H.: Review and annotated bibliography of ancient lake deposits (Precambrian to Pleistocene) in the Western States, U.S. Geol. Surv. Bull., 1080, 1–119, 1964.
Gaiser, E. E. and Johansen, J. R.: Freshwater diatoms from Carolina Bays and other isolated wetlands on the Atlantic coastal plain of South Carolina, U.S.A., with descriptions of seven taxa new to science, Diatom Res., 15, 75–130, 2000.
Hanna, G. D.: Distribution of West American deposits of fossil diatoms, Bios, 27, 227–231, 1956
Héribaud, J.: Les Diatomées Fossiles d'Auvergne, Librairie des Sciences Naturelles, Paris, v–x, 1–166, 1903.
Heribaud, J.: 1920. Diatomees des Travertins D'Auvergne, Imprimerie Medicale et Scientifique, Bruxelles, 206 pp., 1920.
Kelly, M. G.: Short term dynamics of diatoms in an upland stream and implications for monitoring eutrophication, Environ. Pollut., 125, 117–122, 2003.
Kobayasi, H. and Kobayashi, H.: A study of Epithemia amphicephala (Østr.) comb. et stat. nov. and E. reticulata Kütz, With special reference to the areolar occlusion, Proc. Ninth Int. Diatom Symp., Biopress Ltd., Bristol, and Koeltz Scientific Books, Koenigstein, 459–468, 1988.
Kociolek, J. P. and Stoermer, E. F.: Taxonomy, ultrastructure, and distribution of Gomphoneis herculeana, G. eriense and closely related species, Proc. Acad. Nat. Sci. Phila., 140, 24–97, 1988.
Kociolek, J. P. and Van De Vijver, B.: Valve morphology and typification of Epithemia eugeniae W. Smith, a forgotten diatom species described from the Pyrenees Mountains in France (Rhopalodiaceae, Bacillariophyta), Not. Alg., 295, 1–6, 2023.
Kociolek, J. P., Lowe, R. L., Sanchez, K., and Stepanek, J.: 2021. Benthic diatom (Bacillariophyceae) flora of Torch Lake, Michigan, an oligotrophic alkaline ecosystem with evident benthic diatom production, with a consideration of some new and interesting species, Great Lakes Bot., 60, 24–55, 2021.
Kociolek, J. P., Williams, D. M., Hamsher, S. E., Miller, S., and Li, J.: Studies on type material from Kützing's diatom collection VIII. Species assigned to the genera Epithemia Brébisson ex Kützing and Rhopalodia O. Müller, Diatom Res., 39, 159–186, 2024.
Kociolek, J. P., Sala, S. E., Guerrero, J., Uyua, N., Hamsher, S. E., Miller, S., Li, J., and Borsa, T.: Valve ultrastructure, systematics, and diversity of the Rhopalodiales. I. Introduction and consideration of morphological groups within the genus Epithemia Brébisson ex Kützing, Nova Hedw., 120, 109–185, 2025.
Krammer, K.: The Gibberula-group in the genus Rhopalodia O. Müller (Bacillariophyceae). I. Observations on the valve morphology. Nova Hedw., 46, 277–303, 1988a.
Krammer, K.: The Gibberula-group in the genus Rhopalodia O. Müller (Bacillariophyceae). II. Revision of the group and new taxa, Nova Hedw., 47, 159–205, 1988b.
Krammer, K. and Lange-Bertalot, H.: Bacillariophyceae. 2. Teil: Bacillariaceae, Epithemiaceae, Surirellaceae, in: Susswasserflora von Mitteleuropa, edited by: Ettl, H., Gerloff, J., and Mollenhauer, D., Band 2/2, Gustav, Risher, Jena, 1988.
Krebs, W. N. and Bradbury, J. P.: Geologic ranges of lacustrine Actinocyclus species. Western United States, U.S. Geol. Surv. Prof. Pap., 1543-B, 51–73, 1995.
Krebs, W. N., Bradbury, J. P., and Theriot, E. C.: Neogene and Quaternary lacustrine diatom biochronology, western USA, Palaios, 2, 505–513, 1987.
Lange-Bertalot, H. and Krammer, K.: Bacillariaceae Epithemiaceae Surirellaceae. Neae und wenig bekannte Taxa, neae Kombinationen und Synonyme sowie Bemerkungen und Erganzungen zu den Naviculaceae, Bibl. Diatomol., 15, 1–289, 1987.
Meyers, J. D. and Newcomb, R. C.: Geology and ground-water resources of the Swan Lake – Yonna Valleys area, Klamath County, Oregon, U.S. Geol. Surv. open-file report, 151 pp., 1952.
Mills, F. W.: An Index to the genera and species of the Diatomaceae and their synonyms. 1816–1932. Part IX, 607–684, Wheldon & Wesley, London, 1934.
Moore, B. N.: Nonmetallic mineral resources of eastern Oregon, U. S. Geol. Survey Bull., 875, 1–180, 1937.
Müller, O.: Rhopalodia, ein Genus der Bacillariaceen, Bot. Jahr. Syst. Pflanz. Pflanzengeogr., 22, 54–71, 1895.
Müller, O.: Bacillariaceen aus den Natronthälern von El Kab (Ober-Aegypten), Hedw., 38, 274–288, 1899.
Müller, O.: 1900. Bacillariaceen aus den Natronthälern von El Kab (Ober-Aegypten), Hedw., 38, 289–321, 1900.
Newcomb, R. C.: 1958. Yonna Formation of the Klamath River basin, Oregon, Northw. Sci., 32, 41–48, 1958.
Patrick, R. M. and Reimer, C. W.: The diatoms of the United States exclusive of Alaska and Hawaii. Volume 2. Part 2. Entomoneidaceae, Cymbellaceae, Gomphonemaceae, Epithemiaceae, Monographs of the Academy of Natural Sciences of Philadelphia, 13, 213 pp., 1975.
Peterson, N. V. and McIntyre, J. R.: The reconnaissance Geology and Mineral Resources of eastern Klamath County and western Lake County, Oregon, State Oreg. Dept. Geol. Min. Ind., Bull., 66, 70 pp., 1970.
Power, M., Lowe, R., Furey, P. C., Limm, M., Finlay, J., Bode, C., Chang, S., Goodrich, M., and Sculley, J.: 2009. Algal mats and insect emergence in rivers under Mediterranean climates: towards photogrammetric surveillance. Freshw. Biol., 54, 101–115, 2009.
Prescott, G. W. and Dillard, G. E.: 1979. A checklist of algal species reported from Montana 1891 to 1977, Monograph No. 1, Montana Acad. Sci. Suppl. Proc., 38, 1–102, 1979.
Reimer, C. W.: Some aspects of the diatom flora of Cabin Creek raised bog, Randolf Co., Indiana, Proc. Ind. Acad. Sci., 71, 305–319, 1961.
Ruck, E. C., Nakov, T., Alverson, A. J., and Theriot, E. C.: Phylogeny, ecology, morphological evolution, and reclassification of the diatom orders Surirellales and Rhopalodiales, Mol. Phylogen. Evol., 103, 155–171, 2016.
Rushforth, S. R. and Merkley, G. S.: Comprehensive list by habitat of the algae of Utah, USA, Great Bas. Nat., 48, 154–179, 1988.
Servant-Vildary, S., Fourtanier, E., Kociolek, J. P., MacDonald, M. M., and Miette, A.: Nomenclatural issues, types and conserved material from Tempère and Peragallo's Diatomées du Monde Entier, 2nd Edn., Diatom Res., 16, 363–398, 2001.
Sims, P. A.: A taxonomic study of the genus Epithemia with special reference to the type species E. turgida (Ehrenb.) Kütz, Bacillaria, 6, 211–235, 1983.
Skinner, K. G., Dammann, A. A., Swift, R. E., Eyerly, G. B., and Shuck Jr., G. R.: 1944. Diatomites of the Pacific Northwest as filter aids, U. S. Bur. Mines, Bull., 460, 1–96, 1944.
Smith, H. L.: Diatomacearum Species Typicae, No. 152, Boston, Stodder, Centuries I–VI, numbers 1–600; supplement numbers 601–750, 1876.
Smith, W. D.: Diatomaceous earth in Oregon, Econ. Geol., 27, 704–715, 1932.
Stancheva, R., Sheath, R. G., Read, B. A., McArthur, K. D. Schroepfer, C., Kociolek, J. P., and Fetscher, A. E.: Nitrogen-fixing cyanobacteria (free-living and diatom endosymbionts): their use in southern California stream bioassessment, Hydrobiol., 720, 111–127, 2013.
Stoermer, E. F. and Kreis Jr., R. G.: Preliminary checklist of diatoms (Bacillariophyta) from the Laurentian Great Lakes, J. Great Lakes Res., 4, 149–169, 1978.
Stoermer, E. F., Kreis Jr., R. G., and Andresen, N. A.: (1999) Checklist of diatoms from the Laurentian Great Lakes. II, J. Great Lakes Res., 25, 515–566, 1999.
Tempère, J. and Peragallo, H.: Diatomées du Monde Entier, Edition 2, 30 fascicules. Fascicule 2–7. Chez J. Tempère, Arcachon, Gironde, 17–112, 1908.
Tempère, J. and Peragallo, H.: Diatomées du Monde Entier, Edition 2, 30 fascicules. Fascicule 8–12. Chez J. Tempère, Arcachon, Gironde, 113–208, 1909.
Tempère, J. and Peragallo, H.: Diatomées du Monde Entier, Edition 2, 30 fascicules. Fascicule 13–16. Chez J. Tempère, Arcachon, Gironde, 209–256, 1910.
Tempère, J. and Peragallo, H.: Diatomées du Monde Entier, Edition 2, 30 fascicules. Fascicule 17–19. Chez J. Tempère, Arcachon, Gironde, 209–256, 1911.
Tempère, J. and Peragallo, H.: Diatomées du Monde Entier, Edition 2, 30 fascicules. Fascicule 24–28. Chez J. Tempère, Arcachon, Gironde, 353–448, 1913.
Van Landingham, S. L.: Catalogue of the fossil and recent genera and species of diatoms and their synonyms. Part III. Coscinosphaena through Fibula, J. Cramer, Lehre., 1087–1756, 1969.
Vigneshwaran, A., Liu, Y., Kociolek, J. P., and Karthick, B.: A new species of Epithemia (Bacillariophyceae: Rhopalodiales) from the Mula River, Western Ghats, India, with comments on the phylogenetic position of the genera Rhopalodia and Epithemia, Phytotaxa, 489, 171–181, 2021.
Wallace, A. R., Frank, D. G., and Founie, A.: Freshwater diatomite deposits in the western United States, U. S. Geol. Surv. Fact Sheet 2006-3004, 1–2, 2006.