the Creative Commons Attribution 4.0 License.
the Creative Commons Attribution 4.0 License.
 | 26 Jun 2025
| 26 Jun 2025
Analysis of the types of Reinholdella, an important Jurassic foraminiferal genus
Marjorie Apthorpe
The important Jurassic foraminiferal genus Reinholdella is described and redefined based on the extant holotype of the correct type species, Reinholdella dreheri Brotzen, 1948 (= Discorbis dreheri Bartenstein sensu ten Dam and Reinhold, 1942). The invalid neotype of Reinholdella dreheri (Bartenstein), erected by Loeblich and Tappan (1987) (that should have been referred to as the neotype of Discorbis dreheri Bartenstein), is illustrated by scanning electron microscope (SEM) photography and redescribed. Some of the taxonomic confusion surrounding the species Discorbis dreheri Bartenstein, 1937 is illustrated from the variable morphotypes in the type material. An important species ascribed to Reinholdella, R. macfadyeni (ten Dam), is illustrated and its features compared with the type specimen. The holotype of the type species and other species illustrated have only one aperture. Species assigned to Reinholdella require re-assessment.
- Article
(20371 KB) - Full-text XML
-
Supplement
(50954 KB) - BibTeX
- EndNote




The genus Reinholdella Brotzen is of considerable stratigraphic importance in Jurassic foraminiferal assemblages worldwide and includes several species of particular biostratigraphic utility (Gradstein, 1976; Copestake and Johnson, 1989, 2014). However, problems have arisen due to a lack of understanding of the type species morphology; the small size of many species of Reinholdella; the tendency for the critical later chambers to break off, resulting in the absence of the final aperture in most specimens; and the commonly poor preservation or replacement of the originally aragonitic test with pyrite.
The descriptions of the concept of Reinholdella available to the author comprised the illustrations in the Treatise on Invertebrate Paleontology (Loeblich and Tappan, 1964; also 1987), the erroneous designation of the holotype of Discorbis dreheri Bartenstein, 1937 as Reinholdella by Brotzen (1948) (see below), and the brief descriptions and diagrammatic figures by Bartenstein in Bartenstein and Brand (1937). These appeared to present contradictions. In an effort to understand the taxonomy and stratigraphic connotations of Reinholdella in Australian Jurassic material, the author decided to examine the type material.
This process began almost incidentally when, during the course of a short visit to the Senckenberg Museum in Frankfurt in 1995 to examine Jurassic Lageniidae, the author requested to examine whatever material of Discorbis dreheri was available. Several slides of the paratypes of D. dreheri were provided; the holotype was not available without prior arrangement. During the examination of the paratypes, it seemed obvious (from the few specimens mounted with their umbilical sides uppermost) that more than one species was represented.
At the suggestion of Stefan Revets, the author decided that an examination of the types was desirable, and in 2000 Revets facilitated this by borrowing the Brotzen type specimens (F.410) from the Geologische Stichting Haarlem and the Loeblich and Tappan neotype (USNM 383567) from the US National Museum. Both specimen slides were promptly returned to their respective museums after photography. Uncertain at this stage as to what Bartenstein had encompassed within Discorbis dreheri, the author in 1999 requested representative specimens of this species from Heinz Malz (then at the Senckenberg Museum) and in 2000 from Helmut Bartenstein. Some of these specimens survived posting to Australia and were examined as part of this study. These fragile specimens are lodged with the Senckenberg – Research Institute and Natural History Museum, Frankfurt, Germany, under the collection numbers SMF XXVII 14522 to SMF XXVII 14546.
It became obvious during this study that there was a nomenclatural problem that affected the understanding of the entire genus.
The scanning electron microscope allows the examination of small specimens in great detail in a manner impossible 50 years ago when species of the genus were first described. Furthermore, the development of the environmental scanning electron microscope (ESEM™) makes it possible to examine critical small morphological features in type specimens, without coating or otherwise altering the specimen. This paper photographically illustrates the type specimen of the type species of the genus Reinholdella for the first time and redescribes the type species and other species with which it has been linked. It is now possible to more clearly define what is, and what is not, Reinholdella based on the correct type specimen. However, a generic re-assessment of all 29 species presently assigned to Reinholdella is beyond the scope of this paper.
In the course of a discussion on the genus Asterigerina, Brotzen (1948) erected the new genus Reinholdella but without illustrating it and without a full description. Brotzen (1948, p. 126) described the new genus as follows: “This new genus Reinholdella … is characterized by an umbilical and interiomarginal aperture and an inner partition in the chambers as in Lamarckina. This new genus is distinguished from Asterigerina by the shape of the aperture and the situation of the inner partition, from Lamarckina by the lack of an umbilicus and the occurrence of an interiomarginal part of the aperture, from Epistomina by the lack of a lateral aperture”. Brotzen designated the type species as “Asterigerina dreheri, collection Geologische Stichting Haarlem Nr. F.410, original of ten Dam and Reinhold's fig. 1, 1941” (actually, 1942). Ten Dam and Reinhold (1942) had identified this specimen as “Asterigerina dreheri (Bartenstein)”, believing their material to be conspecific with Discorbis dreheri Bartenstein, 1937. Ten Dam and Reinhold reassigned this species to Asterigerina on the basis of the “indistinct angular rhomboid supplementary chambers” on the ventral side.
Brotzen's (1948) citation of the Dutch specimen F.410-1 Asterigerina dreheri as the type species of Reinholdella, with the unstated inference that the species had been described by ten Dam and Reinhold, was corrected in a brief note (Brotzen, 1949). He stated that “The genotype of Reinholdella must be Reinholdella dreheri (Bartenstein) = Discorbis dreheri Bartenstein, 1937. Asterigerina dreheri is only a synonym for Discorbis dreheri Bartenstein”. From this it appears that Brotzen considered that A. dreheri (of ten Dam and Reinhold) and Discorbis dreheri Bartenstein were identical, and he intended Discorbis dreheri Bartenstein to be the type species of his genus. However, this 1949 correction has no taxonomic validity because under the rules of the Code of Zoological Nomenclature, Brotzen would have had to apply formally to the International Commission on Zoological Nomenclature to set aside his mistakes of using the wrong name for the type and of designating the specimen illustrated by ten Dam and Reinhold as the type species; and in its place designating the holotype of Discorbis dreheri Bartenstein as the type species of the genus (ICZN 4th Edition, Article 67.7, Article 72.4.2, and Article 86.3). Because he did not apply to the commission to formally correct his error, his original designation of the Dutch specimen of ten Dam and Reinhold (Asterigerina dreheri (Bartenstein), coll. no. F.410-1 in the Geologische Stichting Haarlem) must stand as the holotype specimen of Reinholdella dreheri Brotzen, the type species of the genus Reinholdella.
To establish the characteristics of the genus Reinholdella, this type specimen (fig. 1 of ten Dam and Reinhold, 1942; see below) is illustrated and redescribed herein.
Two further questions have been raised by the examination of the type specimens by the current author.
2.1 What is Asterigerina dreheri ten Dam and Reinhold?
Ten Dam and Reinhold's (1942) slide F.410 of Asterigerina dreheri contains two specimens, which, based on the current author's examination, appear not to be conspecific. One is recognisably the specimen illustrated in their fig. 1, which Brotzen specifically designated the type specimen of the type species of Reinholdella (specimen designated here F.410 “-1”, described below). The second specimen has a distinctly different morphology and is considered herein to be a separate species from the type. It has been designated specimen F.410 “-2” and is described and illustrated here.
Ten Dam and Reinhold's paper also showed a thin section through the last-formed whorl of Asterigerina dreheri. The very short “toothplates” and small “supplementary chambers” shown on the drawing of this thin section suggest that it is a section of their second species (F.410-2), not of the type species (F.410-1).
2.2 Are Asterigerina dreheri (of ten Dam and Reinhold, 1942) and Discorbis dreheri Bartenstein, 1937 conspecific?
The question of whether the type specimen (F.410-1) is, in fact, the same species as Discorbis dreheri Bartenstein, 1937 is a separate issue from the definition of the type specimen of Reinholdella. Due to Brotzen's formally uncorrected error, Discorbis dreheri Bartenstein was not then, and is not now, the type species of Reinholdella, contrary to the opinion of Loeblich and Tappan (1987, p. 445). In this they followed Brotzen (1948), Hofker (1952), and Ohm (1967). Following the statement of Ohm (1967) that the holotype of Discorbis dreheri is disintegrated, Loeblich and Tappan (1987) designated a neotype for Reinholdella, believing Discorbis dreheri to be conspecific.
This separate question, whether Reinholdella dreheri Brotzen, 1948 (specimen F.410-1) of ten Dam and Reinhold is the same species as Discorbis dreheri Bartenstein, 1937, is discussed below. If the two specimens are not conspecific, then Reinholdella is based on a specimen that has been misidentified by Brotzen. The present author considers that the two species are not the same (see the descriptions below). The new nomenclature used here is required under Articles 11.10 and 67.7 of the Code of Zoological Nomenclature and is discussed in the description under “Remarks” (below).
Much of the above discussion has been summarised in Copestake and Johnson (2014, p. 322) citing the present author as a personal communication. This arose from an earlier 2002 version of this paper that was reviewed by Philip Copestake.
-
Genus Reinholdella Brotzen, 1948
-
Reinholdella dreheri Brotzen, 1948
Plate 1, figs. 1–10; Plate 2, figs. 1–4; Plate 3, fig. 10; Figs. 1–2.
-
1942 Asterigerina dreheri (Bartenstein); ten Dam and Reinhold: 10–11, fig. 1 only.
-
1948 Reinholdella dreheri Brotzen: 126.
-
non 1937 Discorbis dreheri Bartenstein: 192, pl. 6, 45a–c; pl. 8, 42a–d; pl. 10, 47a–d.
-
non 1964 Reinholdella dreheri (Bartenstein); Loeblich and Tappan: C776–777, fig. 637, 2–3.
-
non 1987 Reinholdella dreheri (Bartenstein); Loeblich and Tappan: 445, pl. 476, 13–18.
-
non 2014 Reinholdella dreheri (Bartenstein); Copestake and Johnson: 322, pl. 21, 6, 7, 9.
Material examined. Collection no. F.410 in the Geologische Stichting Haarlem. Specimen designated here F.410-1, in slide containing two specimens.
Locality. Boring Veldzicht, near Winterswijk, the Netherlands, at a depth of 102 and 110 m. Precise location of boring unknown.
Age and stratigraphical level. “Lower Oolitic” = Dogger α= lower Aalenian.
Description. Original description of ten Dam and Reinhold: “Test trochoid, almost planoconvex, dorsal side rather convex, ventral side nearly flat. Periphery subacute. Chambers on the dorsal side regularly coiled, almost 3 whorls, with 6 chambers in the lastformed. Sutures straight, oblique, distinct, limbate, somewhat raised near the center. Ventral side with indistinct angular rhomboid supplementary chambers between the regular series. This charackter is almost invisible at the outside; sections however show clearly these supplementary chambers, which were overlooked by Bartenstein, reason for which he described this species as Discorbis. The center of the ventral side usually shows a low umbilical knob. Sutures more or less indistinct, not raised nor depressed. Wall calcareous with a glassy lustre, finely perforate. Aperture ventral, at the base of the chamber-margin. Diameter: 0.24–0.29 mm. Locality: Proefboring R. 102 and 110 m. The specimens of Bartenstein came mostly from the Dogger of N. W. Germany. Type: Geologische Stichting Haarlem, Coll. No. F 410. Holotype: Senck. Mus. XXVII 666a1” (= Bartenstein's holotype of Discorbis dreheri, now fragmented into three pieces).
Augmented description. Test of moderate size; diameter 0.31 mm; calcareous; trochospiral; biconvex. In edge view the spiral side is more domed and the umbilical side weakly convex. Edge profile subacute where the specimen is intact; most of the youngest periphery of the specimen is broken on the umbilical side, making the profile of the final chamber an unknown factor. In the earlier part of the final whorl, the unbroken periphery is slightly thickened and makes an angle of about 30° with the equatorial plane on the umbilical side and about 40° on the spiral side. The profile rises to a low dome over the axis of coiling on the umbilical side and is uniformly convex on the spiral side. The equatorial outline is subrounded to slightly ovoid. The test consists of the proloculus plus 12 chambers in approximately two whorls, with 6 chambers in the final whorl. On the spiral side, most chambers are visible in the light microscope (Plate 1, figs. 6–7). The proloculus is just visible and is estimated at 35–40 µm across. The second chamber is smaller than the proloculus and barely visible. The six chambers of the inner whorl are rhombohedral, with backward-directed sutures. The six chambers of the outer whorl become progressively more elongated in the direction of coiling, the last two preserved chambers being more than 3 times longer than wide. The final two chambers of the test are broken open, and the spiral surface of these is almost absent, revealing the internal structure.
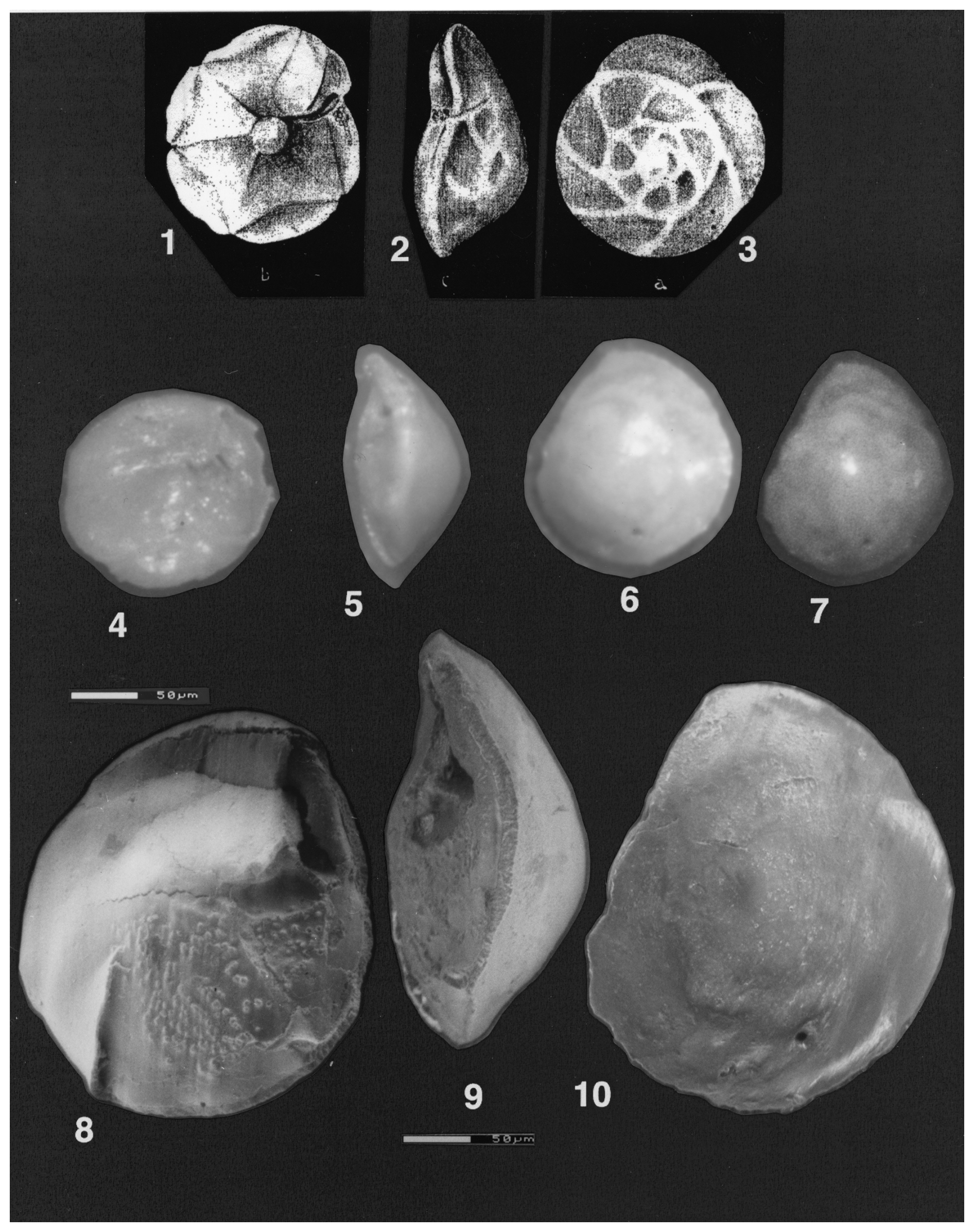
Plate 1(1–10) Reinholdella dreheri Brotzen, 1948 (1–3) Asterigerina dreheri (Bartenstein), fig. 1 of ten Dam and Reinhold (1942); © KNGMG; used with permission. (1) Umbilical view. (2) Edge view. (3) Spiral view. (4–10) Reinholdella dreheri Brotzen, 1948, holotype, specimen F.410-1. (4–7) Light microscope photos. (4) Oblique umbilical view. (5) Edge view. (6) Slightly oblique spiral view. (7) Spiral view. (8–10) ESEM™ photos. (8) Slightly oblique umbilical view. (9) Edge view. (10) Spiral view. Scale bars 50 µm for (8)–(10).
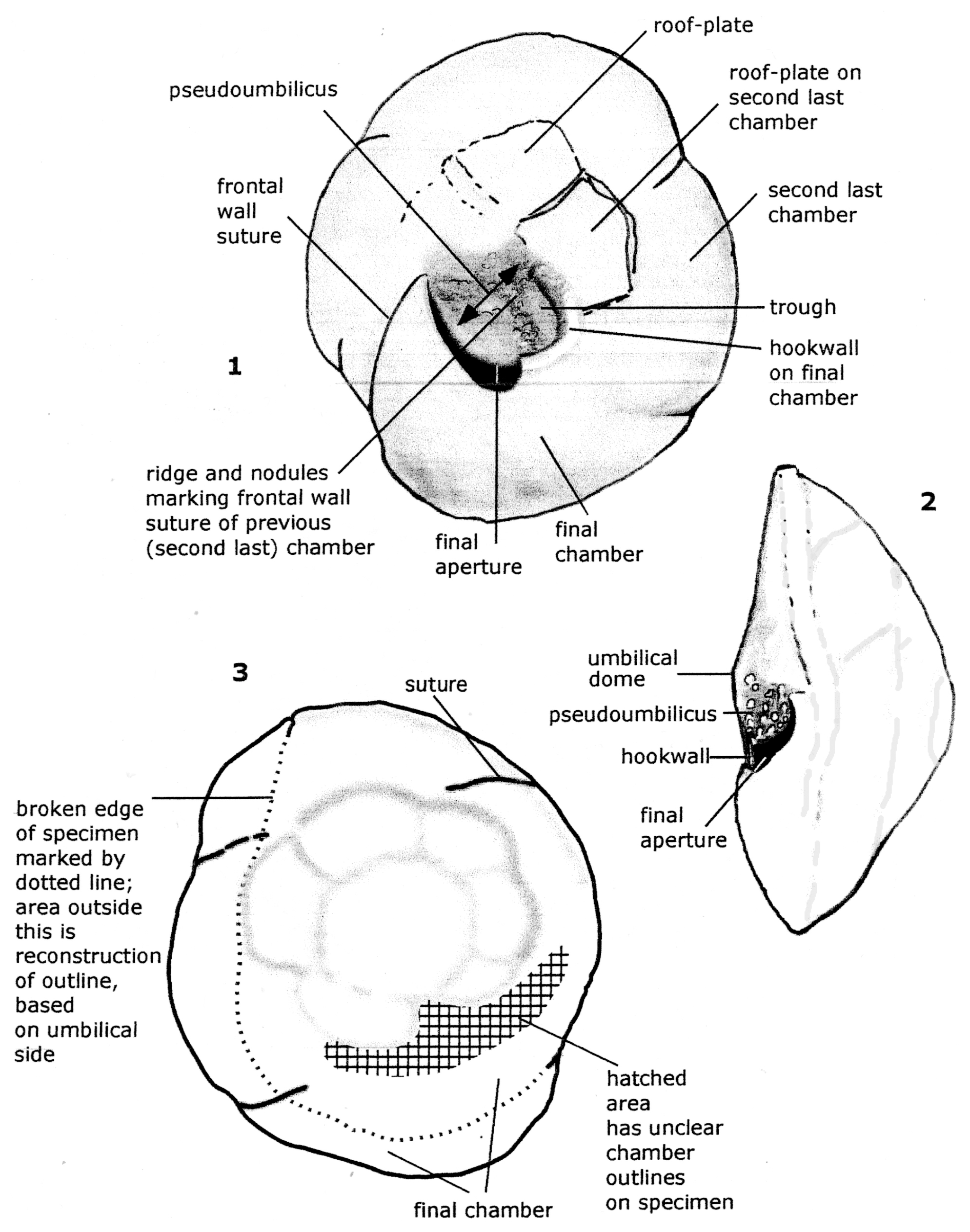
Figure 1Reconstruction of the holotype specimen of Reinholdella dreheri Brotzen, 1948 (F.410-1). Orientation of specimen is identical to that of Plate 1, figs. 8–10.
The septal sutures as seen in the light microscope are oblique and directed backwards; they appear to be flush with the surface in later chambers and only very slightly elevated in some of the earlier chambers. The spiral suture is very slightly raised as a very low broad ridge above the surface. These features are not obvious in the ESEM™. Plate 1, fig. 10 shows the spiral surface photographed at high contrast to try to highlight the low sutural relief on this side of the test. Part of the spiral surface is covered with a layer of glue (at right and bottom edge of photograph, smooth areas with white highlights). However, the impression in the light microscope is that there is almost no sutural relief. The texture of the chamber surfaces is almost entirely smooth. The wall of the final chamber in the ESEM™ appears imperforate (Plate 3, fig. 10); only surface damage of minor pitting and scratches was observed.
The umbilical side is badly damaged, with the last two chambers (0, −1) missing except for the base of walls and wall scars (Plate 1, fig. 8; Plate 2, figs. 1–4; Fig. 2). The third-last (−2) chamber is broken, with the umbilical chamber wall pushed inwards towards the spiral surface, and the umbilical surface is cracked in several places. The foramen and septum bordering this chamber are broken open.
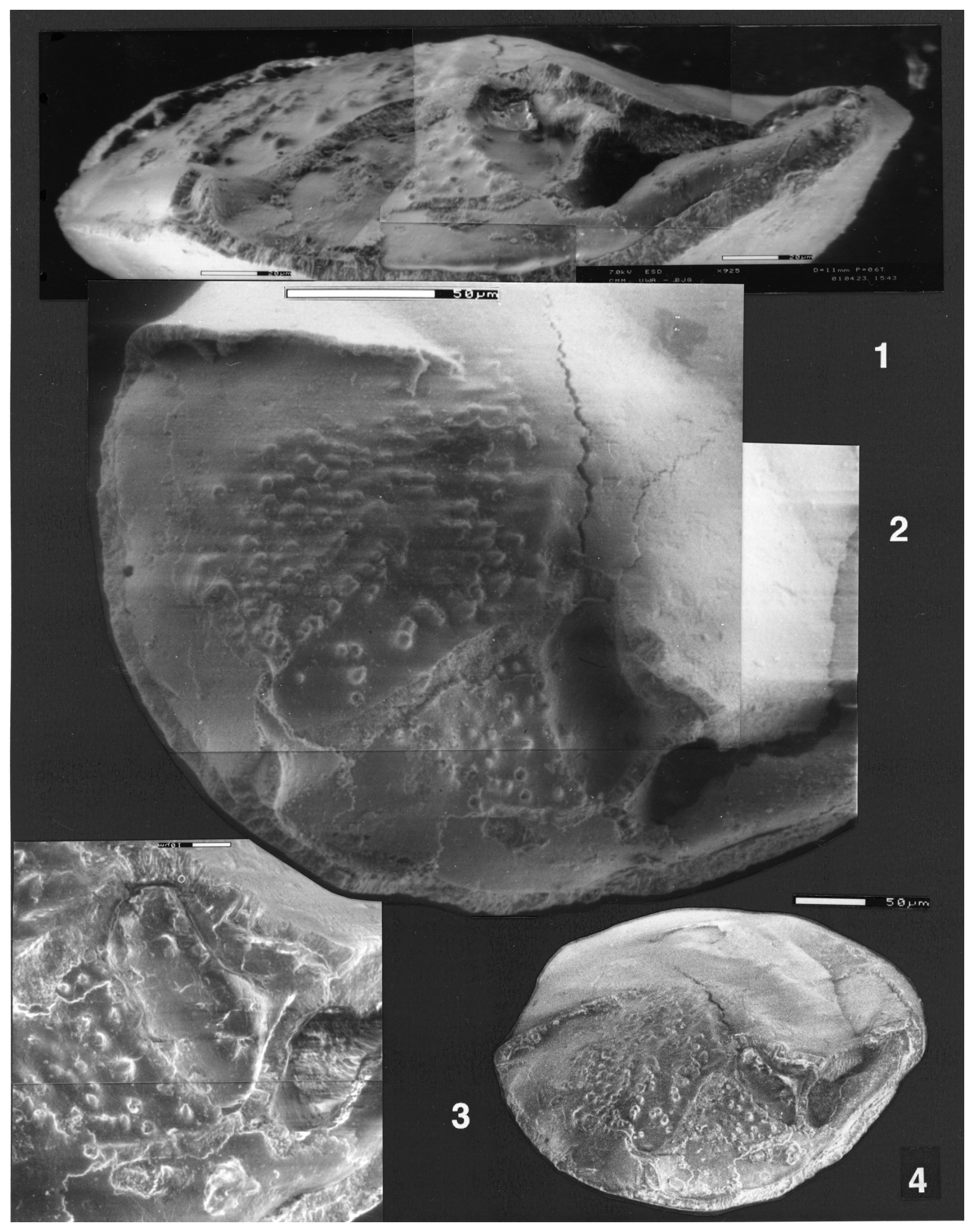
Plate 2(1–4) Reinholdella dreheri Brotzen, 1948, holotype, F.410-1. ESEM™ photographs. (1) Composite partial edge view. Scale bar 20 µm. (2) Composite partial umbilical view, showing the final chamber and penultimate chamber. Scale bar 50 µm. (3) Detail of hookwall, trough, and resorbed frontal wall ridge. Scale bar 10 µm. (4) Oblique umbilical view, whole specimen. Scale bar 50 µm.
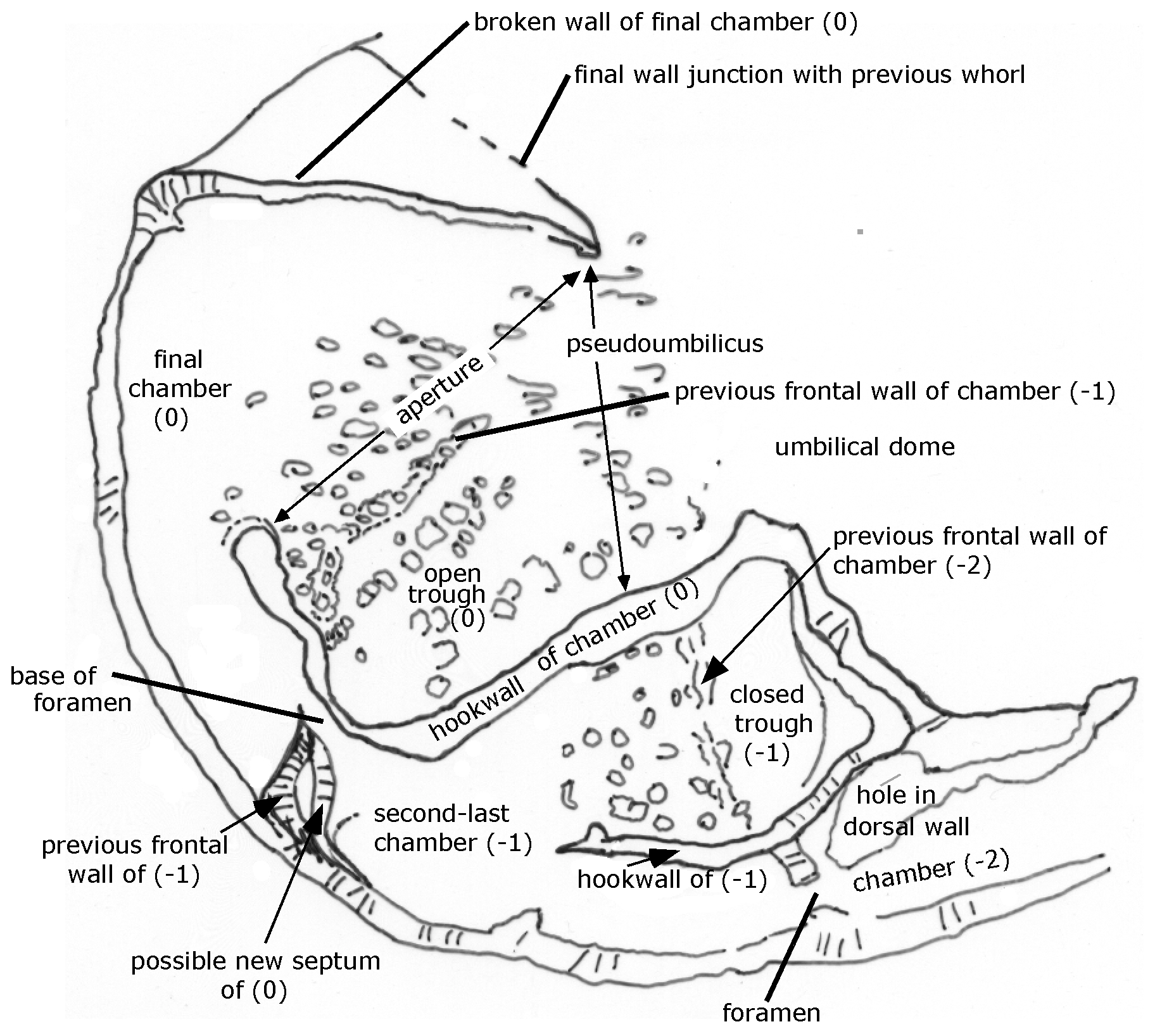
Figure 2Reinholdella dreheri Brotzen, 1948 specimen F.410-1; chamber architecture shown in the last two broken chambers on the umbilical side of the test. Orientation of specimen is identical to that of Plate 2, fig. 2.
The umbilicus in this species is closed (Plate 1, fig. 8; Plate 2, figs. 1–4). The centre of the umbilical side is a low raised dome, which rises smoothly from the surface of chambers −2, −3, and −4. In the ESEM™ there is no visible separation of this axial dome from the surface of these chambers; therefore it does not constitute an umbilical plug. The very prominent umbilical plug shown on the drawing of the specimen (ten Dam and Reinhold, 1942, fig. 1b; Plate 1, fig. 1 herein) is an artefact of the reflection of light off smoother parts of the surface when viewed from certain angles (see, for example, Plate 1, fig. 4). (It is possible that the plug shown by ten Dam and Reinhold is an attempt to combine the features seen in specimen F.410-2, where a shallow groove surrounds part of the central area, with the features of this specimen. However, other features shown in the drawings of their fig. 1 appear to be based on specimen F.410-1.)
The structures of the last two chambers are partly revealed by scars and remnants of the wall on the umbilical side of the test. The chambers do not meet in the centre of the umbilical side. The final chamber (0) frontal wall is present as a broken remnant that attaches to the previous whorl at a low angle. The final wall junction is nearly radial from the periphery, and there is no “primary” apertural opening along the margin of the final chamber or just above the surface of attachment, contrary to the descriptions of Hofker (1952). Thus there are not two apertures, as suggested in many descriptions of Reinholdella (see further below).
The other wall that outlines the final chamber is here termed the “hookwall”. The base of this vertical wall projects radially from the coiling axis for 100 µm and then bends sharply forwards for a further 76 µm, parallel to the periphery. It terminates with a thickened pillar at its free end. This wall is the structure termed the “toothplate” by Hofker (1952) and Ohm (1967) and the “internal pillar-like partition” of Loeblich and Tappan (1964). This structure is neither an internal toothplate (in the sense of Revets, 1993) nor a partition because at the time of its construction it is part of the exterior wall of the chamber (see Figs. 1 and 2). The curve of the final hookwall is visible from the exterior, and it partly delimits the open space (pseudoumbilicus of Hottinger, 2006) into which the aperture faces. (The hookwall does not become an internal structure within the chamber space until it is roofed over by a roof plate during construction of the subsequent chamber; see discussion of chamber construction below.)
There is only one aperture, “interiomarginal” in type. Its position is marked by the gap in wall remnants between the frontal chamber wall and the free end of the hookwall. It faces obliquely backwards, opening into a shallow depressed area that is not axial in position (hence the term pseudoumbilicus). The “aperture” shown in ten Dam and Reinhold's (1942) drawing (Plate 1, fig. 1) is the broken and infolded septum between the penultimate and third-last chambers plus the septal foramen. The edge views shown here on Plate 1, figs. 5 and 9 and Plate 2, fig. 1 are not apertural views. When photographing the specimens, a similar mistake was made as by previous workers, and the specimen was oriented on edge, with the broken chambers facing the viewer. The location of the aperture can be seen in the gap between broken wall scars. A true apertural peripheral view of the specimen would show the aperture to be concealed if the final chamber was intact. The “incorrectness” of this peripheral view was not realised by the author until attempting to draw a reconstruction of specimen F.410-1. By that time the borrowed specimens had been returned.
The frontal wall of any previous chamber is largely resorbed before the addition of the next chamber (Fig. 2). The hookwall of the subsequent chamber is attached behind the position of the previous frontal wall, effectively forming a new back wall for much of the chamber. The remnants of the previous frontal wall are visible as a low ridge and series of tubercules on the floor of the chamber (see Plate 2, figs. 2–4). A small part of the previous frontal wall remains attached to the periphery, forming what must be a short septum. Between this and the hookwall, the intercameral foramen is a resorbed ovoid opening, of which only the base remains here as a smooth surface.
The penultimate and third-last chambers (−1, −2) show the process of chamber formation carried to completion. The frontal wall (as mentioned above) has been resorbed. The chamber is closed off from the pseudoumbilicus by the subsequent vertical hookwall plus a roof plate that arches over the space between the two hookwalls (of 0 and −1). The roof plate over chamber −1 appears as a separate layer in the domed axial area of the specimen and appears to form a continuous layer with the hookwall of the final chamber (0), as far as can be seen from the remnants of these structures, which are broken off (Plate 2, figs. 2–3). In ten Dam and Reinhold's drawing (their fig. 1b; Plate 1, fig. 1 herein), the roof plates (their “rhomboid supplementary chambers”) are shown as a star-shaped outline. However, this outline corresponds to the prominent crack in the umbilical wall of the third-last chamber plus the outer parts of the two hookwalls whose scars are visible in chambers 0 and −1. The indistinct roof plates are actually smaller than the drawing suggests and are extremely difficult to see in the light microscope. Only 2.5 roof plates actually survive on the specimen, over the first three chambers in the final whorl. The roof plate over the penultimate chamber has been broken away to show the pseudoumbilicus beneath (= “supplementary chamber”) (Plate 2, figs. 2–3). (The final chamber (0) should never have a roof plate.) The roof plates are not visible as separate structures in the ESEM™.
Remarks. The nomenclature given here is required under Article 11.10 and Article 67.7 of the International Code of Zoological Nomenclature (ICZN, 1999). Brotzen expressly used the subsequent identification by ten Dam and Reinhold rather than the original Discorbis dreheri Bartenstein, thereby establishing the new nominal species Reinholdella dreheri Brotzen, 1948 for the Dutch taxon actually involved. That Brotzen did not consider (or did not realise) that this was a mistake does not alter the situation. Brotzen's (1949) subsequent attempt to remedy his perceived mistake in attributing his type species to the wrong authorship has been mentioned above (see “Historical background and nature of the problem” section). Brotzen also misquoted (as “1941”) the date of publication by ten Dam and Reinhold; the correct date is 1942.
Thus the nomenclature of this specimen can be summarised as follows: Reinholdella dreheri Brotzen, 1948, type species of Reinholdella now fixed herein (under Article 70.3 of the Code of Zoological Nomenclature), identified as Asterigerina dreheri (Bartenstein, 1937) by ten Dam and Reinhold (1942); subsequently designated by Brotzen as the type species of Reinholdella; under Article 11.10, now deemed to have been erected by Brotzen as the nominal species Reinholdella dreheri Brotzen, 1948.
3.1 Description of a second specimen in the type slide (F.410-2)
-
Reinholdella sp.
Plate 3; figs. 1–9; Plate 4, figs. 1–2; Fig. 3.
-
1942 Asterigerina dreheri (Bartenstein); ten Dam and Reinhold: 10–11, fig. 2?
-
2014 ?Reinholdella dreheri (Bartenstein); Copestake and Johnson: 322–323, plate 21, figs. 6, 7, 9.
Material examined. Collection no. F.410 in the Geologische Stichting Haarlem; specimen no. 2 in slide, designated here F.410-2.
Description. Test calcareous; trochospiral; of moderate size; diameter 0.312 mm; outline circular. Profile in edge view unequally biconvex, approaching a planoconvex shape, the spiral side being strongly domed and the umbilical side very weakly convex. Edge profile subacute to acute, with a thickened rim developed around the periphery on the earlier chambers of the last whorl.
The test consists of 17 chambers plus the proloculus, arranged in three whorls. The highly domed spiral side shows, faintly, the proloculus plus about five chambers in the innermost whorl. These chambers are rhombohedral and slightly longer (in the direction of coiling) than wide (Plate 3, figs. 3, 7). The surface over this whorl is smoothed and partly obscured by secondary lamination. There are six chambers in the second whorl, longer than wide, rhombohedral to crescentic, and with the sutures thickened and slightly raised. The third whorl consists of six chambers, very long, crescentic, and narrow, sloping steeply to the periphery and with thickened, slightly raised oblique sutures. The outline of the final whorl is slightly petaloid. The chamber arrangement and sutures on this side are seen more clearly in the light microscope than in the ESEM™, in which some details are obscured by glue on the spiral surface. The surfaces of the later chambers are light brown, and the thickened sutures are white to cream in colour. The spiral surface of the later chambers may be very finely perforate (see Plate 3, fig. 9), but the surface texture is mostly obscured by surface damage and patches of glue.
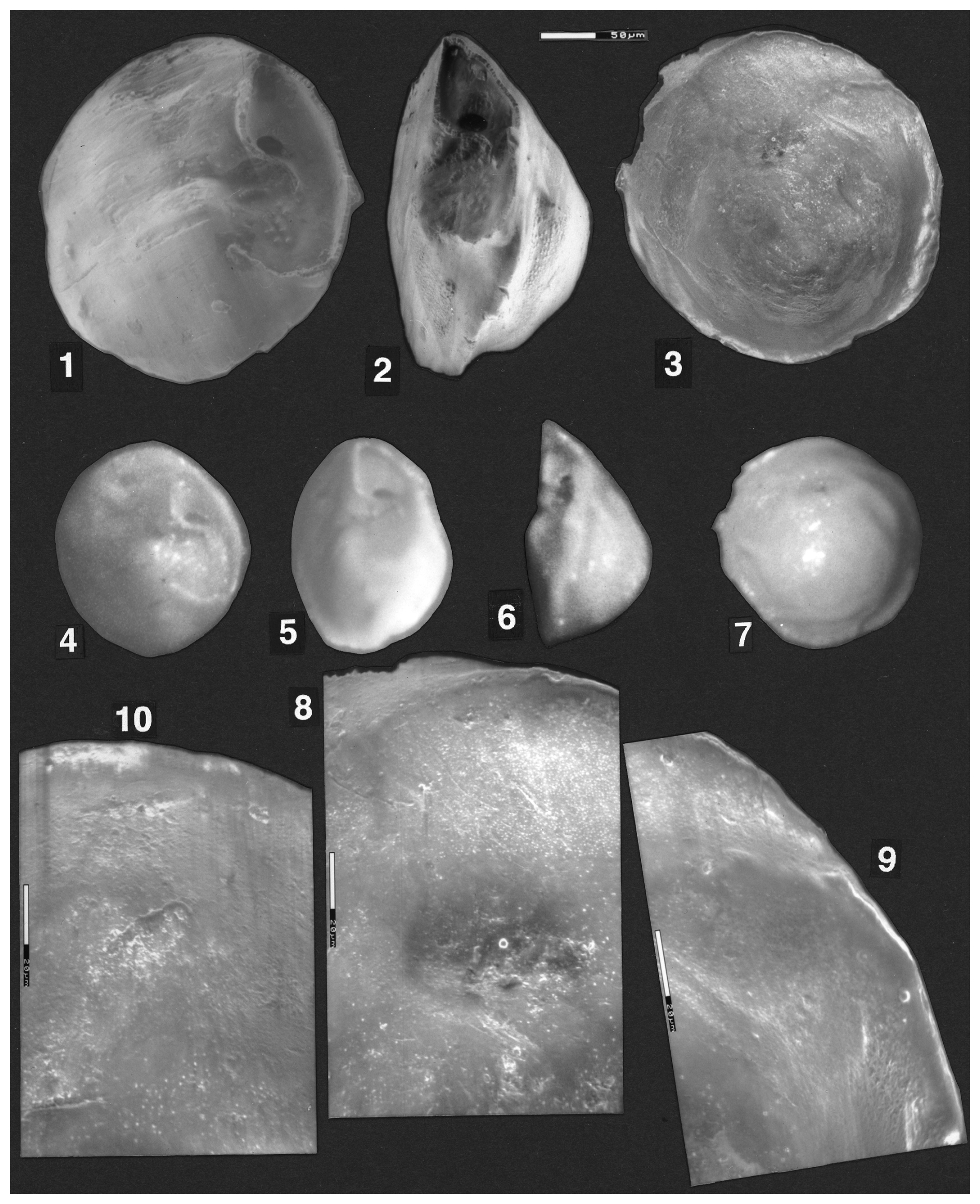
Plate 3(1–9) Reinholdella sp. F.410-2. (1–3) ESEM™ photographs; scale bar 50 µm. (1) Oblique umbilical view. (2) Edge view. (3) Spiral view. (4–7) Light microscope photographs. (4) Umbilical view. (5) Oblique umbilical/edge view. (6) Edge view. (7) Spiral view. (8–9) ESEM™ photos. Spiral surface of wall of last two preserved chambers. Wall appears smooth in some areas and finely perforate in others. Scale bars 20 µm. (10) Reinholdella dreheri Brotzen, 1948, holotype, F.410-1. Detail of last-preserved chamber wall on spiral side.
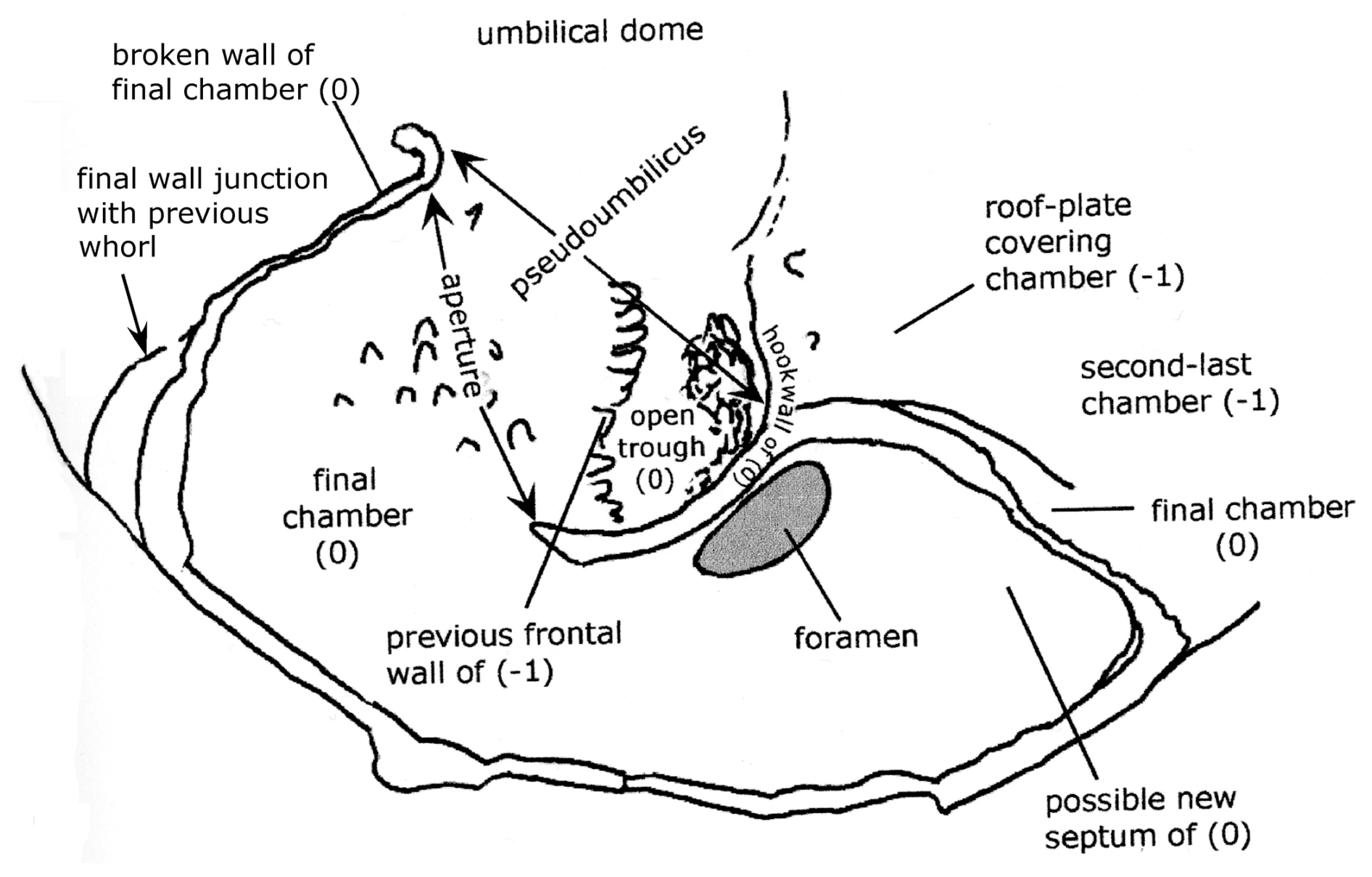
Figure 3Reinholdella sp. specimen F.410-2. Chamber architecture of final chamber on the umbilical side. Orientation of specimen as for Plate 4, fig. 2.
The umbilical surface is smooth, appearing imperforate. The peripheral outline is slightly lobulate. The umbilical side of the specimen is very weakly convex. The umbilicus is closed, with the centre of the umbilical side consisting of a raised thickened area. This raised area is separated from the penultimate chamber by a shallow depression and thus resembles an umbilical plug when viewed from certain angles (Plate 3, figs. 1, 4–6); but when seen from the periphery near the final chamber, it appears to be continuous with the chamber surface of older chambers (Plate 4). The final chamber is broken off. The frontal wall is more curved than in specimen F.410-1. The suture between the last two chambers on the umbilical side is clearly depressed. Most importantly, the hookwall is much shorter and is gently curved throughout its length. Correspondingly, the septum is larger than in F.410-1, extending half way towards the centre and forming a concave surface towards the final chamber (Plate 4, fig. 2). The septum lies behind the position of the previous convex frontal wall; consequently, the septum is interpreted as a “new” structure, not forming part of the previous frontal wall. The intercameral foramen is an oval opening in the new septum on the peripheral side of the hookwall (Plate 4, fig. 2; Fig. 3). It is the result of resorption and does not correspond in position with the aperture. The resorbed previous frontal wall (of chamber −2) is present as a low ridge. The final aperture is indicated by the gap between the end of the hookwall and the end of the frontal wall. The aperture faces obliquely backwards into a narrow open pseudoumbilicus formed by the curve of the hookwall. The floor of the apertural opening contains a number of tubercules, formed by the new inner lamella of the last chamber (Plate 4, figs. 1–2).
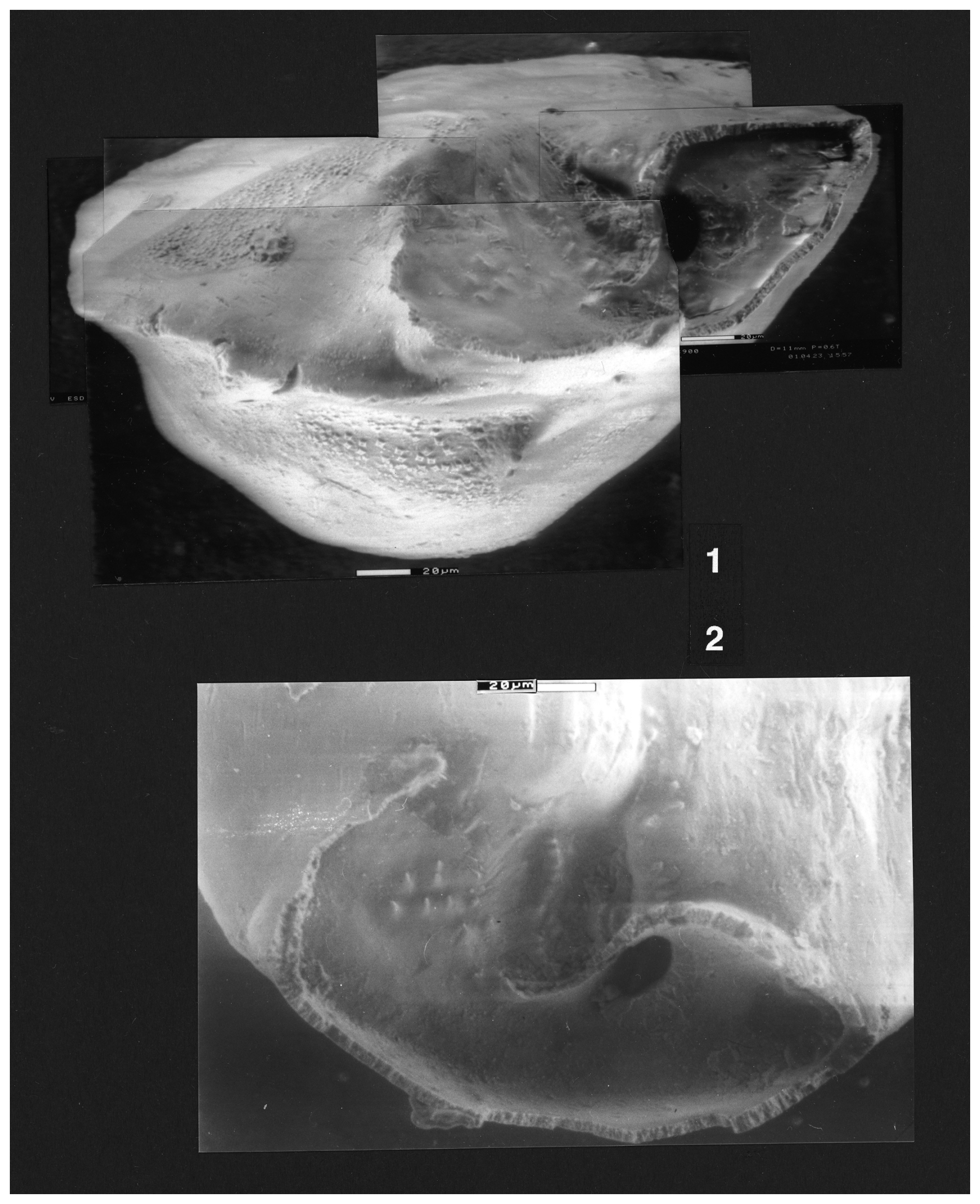
Plate 4(1–2) Reinholdella sp. F.410-2. (1–2) ESEM™ photographs. (1) Oblique edge view (not apertural view) looking across the broken remnants of the final chamber, with the apertural gap in the centre and the intercameral foramen the black oval to the right. (2) Umbilical view of broken final chamber, with frontal wall to left, apertural gap in centre, and secondary intercameral foramen the dark oval at the right. Scale bars 20 µm.
Ten Dam and Reinhold's (1942) fig. 2 is a thin section through the last-formed whorl of a broken specimen, probably of this second species of Reinholdella. The hookwalls shown in this section are very short and consequently close to the axis of coiling, as in this specimen (F.410-2).
In summary, this specimen differs from F.410-1 by its more unequally biconvex test; the raised sutures on the spiral side; and in its shorter, more curved hookwall. The presence of only one specimen of each form in the type slide makes further comparison impossible at present. However, it appears that this second specimen is not part of the type species of Reinholdella (unless Reinholdella dreheri Brotzen, 1948 is found in future to have a wide range of variability which encompasses the differences noted above). It is therefore left in open nomenclature, as only one specimen of the form is presently available for this study. The locality of the borehole from which the specimens came is presently unknown (according to the curator, collection Geologische Stichting Haarlem, written communication to Stefan Revets), a factor that prevents the analysis of other specimens.
The question of whether Reinholdella dreheri Brotzen, 1948 (= Asterigerina dreheri of ten Dam and Reinhold) is conspecific with Discorbis dreheri Bartenstein, 1937 hinges on the question of the morphology of the latter species, which is explored in some detail in this paper. As has been pointed out by previous authors (Ohm, 1967; Loeblich and Tappan, 1987), the holotype of Discorbis dreheri Bartenstein is almost destroyed and consists of three pieces (Heinz Malz, personal communication, 1999) either due to the expansion of marcasite inside the specimen or due to dissolution caused by acidification of the mounting medium, gum tragacanth. Some of the paratypes have also disintegrated by the same process. The extant paratypes represent at least two separate species, one of which does not correspond to the description of the species given by Bartenstein and Brand (1937). Some of these paratypes were examined in the Senckenberg Museum by the author, and sketches are shown herein (see below; Fig. 4).
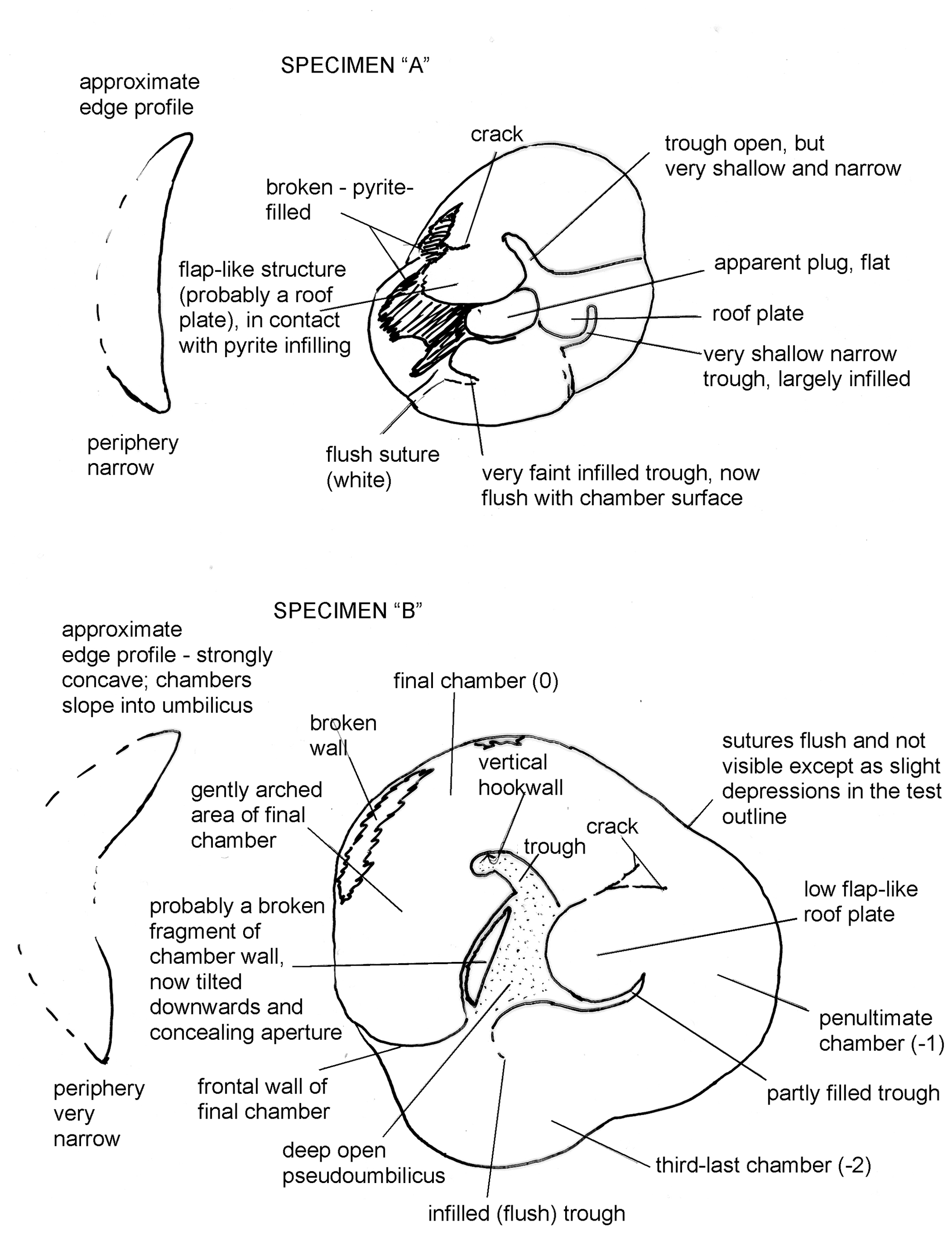
Figure 4(a, b) Sketches of two paratypes of Discorbis dreheri Bartenstein, 1937. Both are un-numbered specimens in Senckenberg Museum assemblage slide XXVII 666 a 2–5.
Due to the condition of the holotype, Loeblich and Tappan (1987) attempted to stabilise the concept of Reinholdella by designating a neotype for Discorbis dreheri Bartenstein, 1937. This specimen (USNM no. 383567) was selected from “topotypes” sent to them by Bartenstein. Unfortunately this neotypification is invalid for three reasons:
-
As indicated previously, the type specimen of the type species of Reinholdella Brotzen is the Dutch specimen F.410 (= Asterigerina dreheri, fig. 1 of ten Dam and Reinhold, 1942), and that specimen (F.410-1) is extant. The type specimen was thus incorrectly cited by Loeblich and Tappan, following Brotzen (1949).
-
While the holotype of Discorbis dreheri Bartenstein, 1937 may be “unrecognizable” (Loeblich and Tappan, 1987, p. 445), it is not totally destroyed, being now in three pieces, and therefore erection of a neotype for that species, however desirable, is not permitted under ICZN Article 75.1 (fide Heinz Malz and Stefan Revets) without a formal application to set aside the holotype (Article 75.5).
-
Loeblich and Tappan's drawing and description of their neotype is incorrect in critical features. It does not accurately represent the apertural characteristics of the specimen (which is broken). Nor does it represent the morphology of Reinholdella Brotzen, which has a closed umbilicus (see above). The specimen itself does not represent Discorbis dreheri Bartenstein (which has an umbilical plug: see below). However, because of the wide circulation achieved by the publication of this illustration in the Treatise on Invertebrate Paleontology, Loeblich and Tappan's neotype specimen has been illustrated here by ESEM™ and the specimen redescribed in detail.
Due to the historical importance of Discorbis dreheri in the definition of Reinholdella, its morphology is examined here using some of the paratypes in the Senckenberg Museum, the neotype designated by Loeblich and Tappan, and then further specimens from the type sample of D. dreheri, supplied by Bartenstein and Malz. These latter specimens of D. dreheri have been lodged in the Senckenberg Museum, Frankfurt, Germany, under the catalogue numbers SMF XXVII 14522 to SMF XXVII 14546.
4.1 Description of Discorbis dreheri Bartenstein, 1937
-
Discorbis dreheri Bartenstein, 1937
-
1937 Discorbis dreheri Bartenstein, in Bartenstein and Brand, 1937: 192, pl. 6, fig. 45 a–c; pl. 8, fig. 42a–d; pl. 10, fig. 47a–d.
Material examined. Paratypes: Bartenstein and Brand, 1937: plate 8, fig. 42 b–d : Senckenberg Museum XXVII 666 a 2–5; plate 10, fig. 47a–d: Senckenberg Museum XXVII 666 a 6–8.
Additional specimens from the type locality, loaned from the Senckenberg collection, sent by Malz and Bartenstein: Senckenberg Museum SMF XXVII 14522–SMF XXVII 14546.
Material not examined. Holotype: Bartenstein and Brand, 1937: plate 8, fig. 42a: Senckenberg Museum XXVII 666 a1. Spiral side only illustrated. Specimen now broken and unrecognisable (fide Ohm, 1967; Heinz Malz, personal communication, 2000).
Paratypes: Bartenstein and Brand, 1937: plate 6, fig. 45a–c: Senckenberg Museum XXVII 666 b 3–4.
Type locality. Hambühren WA 2, 342 m.
Description. Translated from Bartenstein and Brand (1937, p. 192):
“Size of test: Diameter 0.24 mm.
Diagnosis: Test of the genus Discorbis with flat or concave umbilical side, convex spiral side, and aperture situated at the umbilical margin.
Description: Test circular-round, spiral side dome-shaped, umbilical side flat or weakly concave, in the middle with a button-shaped thickening. Sutures flat or slightly depressed, but in the middle of the spiral side (of 2–3 whorls), as ridges, mostly raised. Test surface glassy, finely porous. Aperture with a well defined split, or strongly lobed, on the umbilical side of the youngest chamber.
Occurrence: The new species is found not rarely in the Lias ζ [= Middle–Upper Toarcian], Dogger α (opalinus zone) [= lower Aalenian] and β (ludwigia zone) [= middle Aalenian].”
Remarks. Bartenstein illustrated his new species Discorbis dreheri only on the spiral side of the holotype and on the spiral side of eight paratypes. Only two paratypes were shown on the umbilical side, where the characteristic features of the genus and species are located. There are no figures given of the species in edge view. Thus some key features of the holotype are not known at present. The species description does not state whether the periphery of the holotype of Discorbis dreheri was acute or narrowly rounded.
In a letter to the author dated 30 April 2001, Bartenstein stated “In 1937 Discorbis dreheri was introduced as a new species for which the umbilical knob [Bartenstein's emphasis] is a characteristic feature for further determinations”.
4.2 Some paratypes of Discorbis dreheri Bartenstein
Material examined. Paratypes: Bartenstein and Brand, 1937: plate 8, fig. 42b–d: Senckenberg Museum XXVII 666 a 2–5; plate 10, fig. 47a–d: Senckenberg Museum XXVII 666 a 6–8.
Material not examined. Paratypes: Bartenstein and Brand, 1937: plate 6, fig. 45a–c: Senckenberg Museum XXVII 666 b 3–4.
Remarks. This investigation examined some of the paratypes of Discorbis dreheri Bartenstein in the Senckenberg Museum in Frankfurt. These paratypes represent at least two species (see Fig. 4). This is perhaps the reason that Ohm (1967) did not designate a neotype from among the surviving paratypes.
The specimens examined were grouped with a number of specimens in each slide, and the slides were numbered XXVII 666 a 2–5 and XXVII 666 a 6–8. (The slide labelled XXVII 666 b 1–4 was not seen due to a lack of time.) It was not possible to relate particular numbers to individual specimens nor in most cases to the illustrations of Bartenstein and Brand (1937). Of the seven paratypes seen in two slides, four specimens had disintegrated. The three surviving specimens appeared to be too fragile to turn over. Thus it is not known which of the illustrated specimens survived and were examined. As Ohm (1967) commented, those paratypes examined belong to at least two different species. Neither of the intact species appears identical to the Loeblich and Tappan neotype described below or to the type specimen of Reinholdella described above. One paratype (designated A) is a concavo-convex form, with four or possibly five chambers in the final whorl and a flat plug in the umbilicus (Fig. 4a). This plug appears to be distinctly separate from the roof plates and corresponds to Bartenstein and Brand's (1937) description of a “button-shaped thickening”. This specimen appears to correspond to the description given of the species. The final chamber on this specimen is broken away. The other intact paratype (designated B) illustrated in Fig. 4b has three chambers in the final whorl and narrowly rounded margins. It is unlike the other paratype and differs from the various concavo-convex forms that are present in the type sample (Hambühren WA 2, 342 m) specimens sent to me.
Description of paratype Specimen “A” (unnumbered specimen in Senckenberg Museum assemblage slide XXVII 666 a 2–5). The specimen was viewed from the flat umbilical side only. The profile is plano-convex to slightly concavo-convex (see Fig. 4a). The umbilical side with an ovoid, lobulate outline has four chambers visible in the final whorl. The last-preserved chamber occupies just over one-third of the periphery; it is not certain whether an additional chamber was present, which may have been broken off. Near the peripheral edge, this last chamber is broken open, revealing an infilling of pyrite or marcasite within the chamber. There is also an area of pyrite or marcasite largely covering the fourth-last chamber. The last chamber is only slightly inflated. It has a large flap-like area, similar to the roof plate, projecting almost to the centre of the specimen. The pseudoumbilicus between the hookwall and this flap-like area is open but is very shallow and narrow. No sign of the aperture could be seen, although it might be present underneath the flap-like area. However, it is possible that this flap is actually a secondary roof plate and that this specimen may have the final chamber and its open aperture broken away. Due to the very small size of the specimen, electron microscope examination would be needed to clarify this point. The suture between the last two preserved chambers is depressed. The sutures between the earlier chambers become progressively thickened by secondary lamination. The suture between the third-last and fourth-last chambers is white and flush with the chamber surface. The penultimate preserved chamber has a very shallow, narrow trough between the roof plate and the peripheral area of the chamber (Fig. 4a). The third-last chamber has a trough now completely infilled by secondary lamination, and this area is therefore flush with the chamber surface. However, the area of the trough is very faintly visible in the light microscope as a different colour of wall. The umbilical area is closed by what appears to be a plug, essentially level with the chamber surface. At first the plug was thought to be a roof plate attached to the third-last chamber, but a very faint, filled trough occurs closer to the periphery on this chamber, and the position of the roof plate can be suggested. It appears to be separate from the central plug.
Description of paratype Specimen “B” (unnumbered specimen in Senckenberg Museum assemblage slide XXVII 666 a2–5).
The specimen was seen only from the umbilical side but appears to be fairly strongly convex on the spiral side and is concave on the umbilical side (see Fig. 4b). The periphery appears to be sub-acute or more narrowly rounded than Specimen “A”. The test outline is trilobate. The umbilicus is open and deep. A small amount of pyrite/marcasite obscures the base of the umbilicus. Three chambers are visible on the umbilical side. The last one occupies nearly half of the periphery. It appears to be the final chamber of the specimen in life. It is unbroken at the frontal suture between the periphery and the umbilicus, that is with no apertural opening or foramen on this part of the chamber face. Along the peripheral edge there is a broken area of wall, forming a jagged slit. The chamber is gently arched and slopes from the periphery downwards into the umbilical hollow. The pseudoumbilicus and the hookwall are clearly visible. The hookwall is vertical. The aperture opening could not be observed directly because a broken fragment of chamber projects downwards into the apertural opening (see Fig. 4b). The penultimate chamber is separated from the final chamber by a slight indentation in the peripheral outline. The last suture is almost flush with the chamber surfaces. A flat, low flap, bounded by the last suture and the partly filled trough of the second-last chamber, projects into the umbilicus. This flap is interpreted as the secondary roof plate, which is inferred to have covered the aperture and partly infilled the trough. The third-last chamber likewise can be discerned by the lobate outline of the periphery. The suture between this and the following chamber is flush with the surface. The trough of this chamber has likewise been infilled by secondary lamination and is now flush and only very faintly visible in the light microscope. The flap (interpreted as the roof plate) covering this trough projects as a slight bulge into the open umbilicus.
The two specimens described here belong to different species and possibly genera. Only Specimen “A” appears to correspond to the description of Discorbis dreheri as having a “button-shaped thickening” in the middle of the umbilical side. Specimen “B” is a trilobate, three-chambered form with an open umbilicus and flush sutures. It does not appear to be referable to Discorbis dreheri.
4.3 The neotype of Discorbis dreheri Bartenstein designated by Loeblich and Tappan (1987)
-
Discorbis dreheri Bartenstein, 1937
Plate 5, figs. 1–7; Fig. 5.
-
1937 Discorbis dreheri Bartenstein, in Bartenstein and Brand, 1937: 192, plate 6, fig. 45a–c; plate 8, fig. 42a–d; plate 10, fig. 47a–d.
-
1964 Reinholdella dreheri (Bartenstein); Loeblich and Tappan: C776–777, fig. 637: 2a–2c (reproduced herein, with permission).
-
1987 Reinholdella dreheri (Bartenstein); Loeblich and Tappan: 445, plate 476, figs. 13–15 (Neotype, USNM 383567, designated by Loeblich and Tappan).
-
non Reinholdella dreheri Brotzen, 1948.
Material examined. USNM 383567, borrowed from the Smithsonian Institution, Washington, D.C. Specimen sent to USNM by Helmut Bartenstein, from Hambühren WA 2 at 342 m; opalinum zone (Aalenian).
Description. Test small, calcareous, free, low trochospiral, plano-convex. Dimensions as seen in the ESEM™: length 174 µm; width 140 µm; thickness 79 µm (plate 5, figs. 1, 3, 4). All 12–13 chambers are visible in the two whorls on the spiral side. The final chamber is broken off. The initial chamber is very hard to see due to secondary lamination. Only 4.5 chambers of the final whorl are visible on the umbilical side (Plate 5, figs. 4, 7). The broken final chamber exposes the fifth-last chamber. The peripheral outline as seen from the spiral and umbilical sides is ovoid and slightly lobulate.
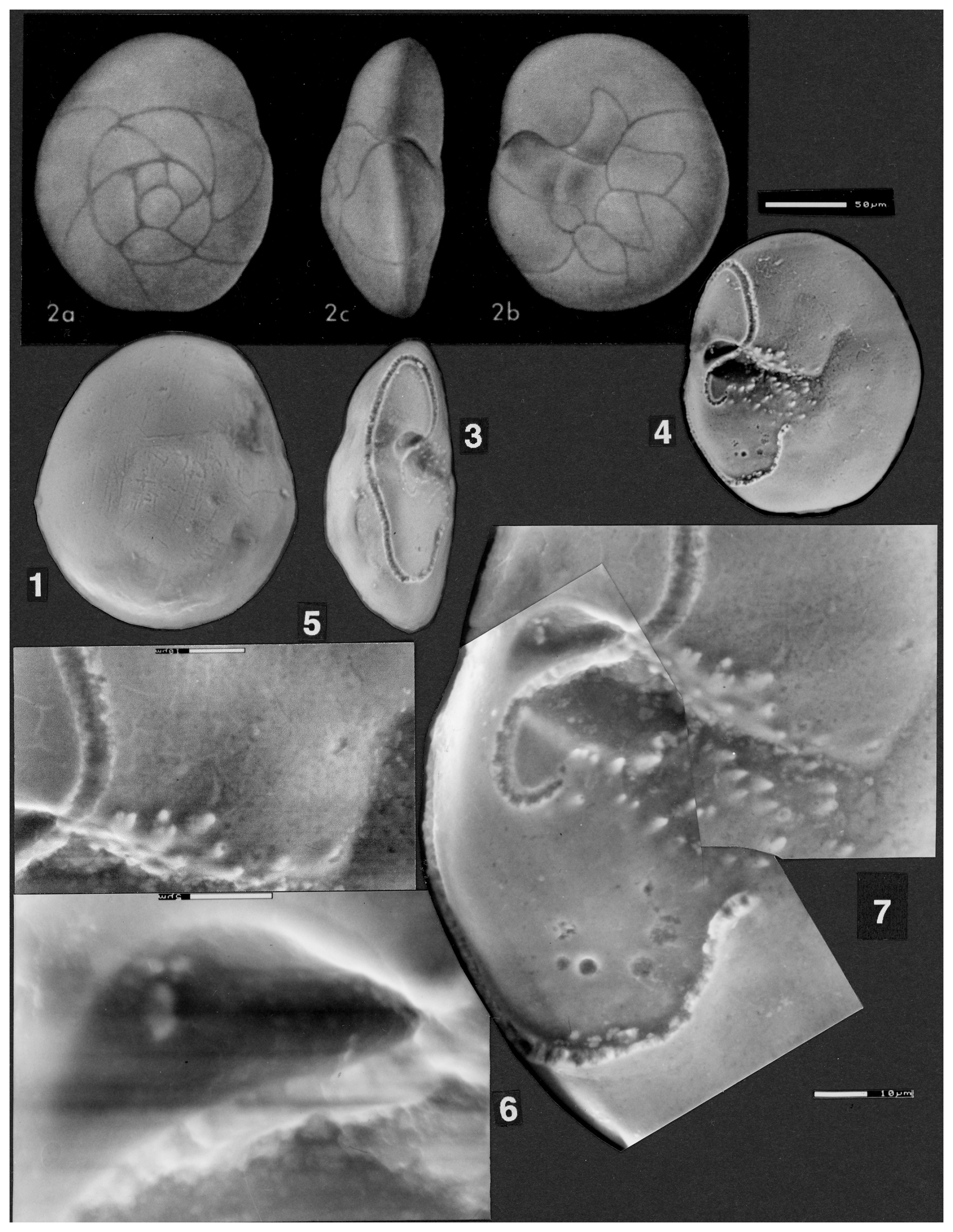
Plate 5(1–7) Discorbis dreheri Bartenstein, 1937, USNM no. 383567; neotype of Reinholdella designated by Loeblich and Tappan (1987). (2a, 2b, 2c) Illustration of specimen from Loeblich and Tappan (1964, fig. 637, 2a–c); © Geological Society of America, used with permission. (1) ESEM™, spiral view. (3) ESEM™, edge view, showing the broken-off final chamber. (4) ESEM™, slightly oblique umbilical view. The apertural opening is clearly visible between the hookwall and the final chamber wall. The intercameral foramen lies above the hookwall. Scale bar 50 µm for figs. 1–4. (5) ESEM™ of the final roof plate showing the apparently imperforate surface, with minor etching. Scale bar 10 µm. (6) ESEM™ of the arched intercameral foramen, which in this specimen is closed by what appears to be shell wall material. Scale bar 5 µm. See also (3) and (7). (7) ESEM™ (composite) of the area of the final chamber, showing in more detail the umbilical position of the only aperture, the hookwall with the resorbed intercameral foramen above it, and the closed trough below, as well as the roof plate which has closed off the aperture of the previous chamber. Scale bar 10 µm.
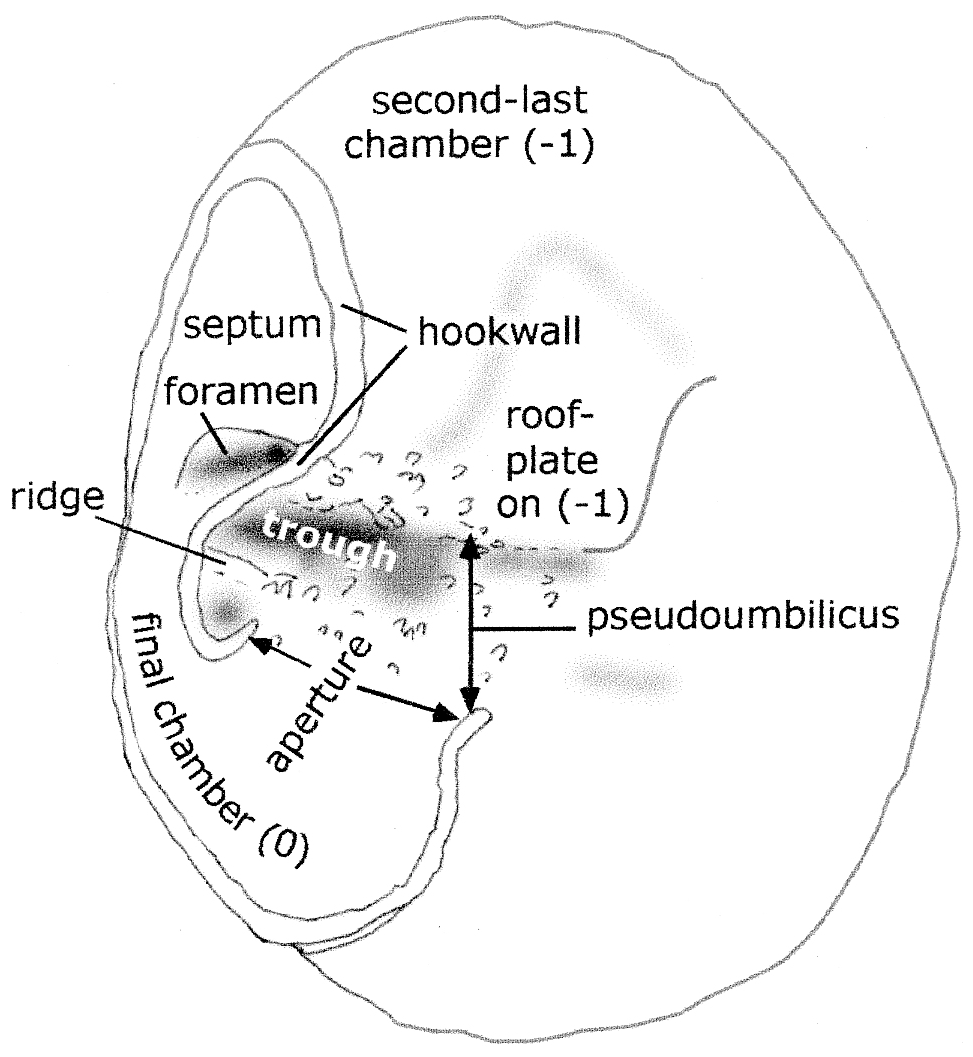
Figure 5Discorbis dreheri Bartenstein, 1937: neotype designated by Loeblich and Tappan (1987), specimen USNM 383567. Features of chamber morphology on umbilical side, as shown on Plate 5, figs. 4 and 7.
In edge view, the peripheral margin is narrowly rounded. No keel is present. There may be slight additional thickening of the periphery on the umbilical side due to secondary lamination, but this could not be seen clearly in the specimen.
On the spiral side, the spiral suture between the whorls is slightly but distinctly depressed. The intercameral sutures on the spiral side are oblique to the spiral suture; near the periphery they are flush or almost flush with the chamber surface; approaching the spiral suture they become slightly depressed (Plate 5, fig. 1).
On the umbilical side the intercameral sutures are radial and slightly depressed in younger chambers. The sutures become flush in older chambers due to secondary lamination.
The wall is shiny and smooth on both sides and is considered to be imperforate throughout, including the roof plates (Plate 5, fig. 5). No regular pores could be seen on either the spiral or umbilical side of the test. At extreme magnifications, minor specimen damage and irregular, elongate etch marks 1–2 µm in size are discernible on the umbilical side on exterior chamber walls. Within the floor of the broken final chamber, two circular holes and four smaller pits are visible (see Plate 5, fig. 7), but these are on the inner lamella of the chamber that abuts the previous whorl and cannot be considered as pores in the (exterior) wall. No pores were observed on the secondary roof plates, which form a star-shaped ring around the umbilicus. A few scattered, irregular pits on the last two roof plates, ranging from 1 to 3 µm across, are regarded as being due to slight etching or damage to the wall, as these depressions are extremely shallow (Plate 5, figs. 4, 5, 7).
The pseudoumbilicus is shallow and open only in front of the final chamber. The pseudoumbilicus adjacent to older chambers is secondarily closed by the addition of the large roof plates. The portion of the pseudoumbilicus remaining open is asymmetrical and consists partly of a narrow and deeper depression, here termed the trough (see Fig. 5).
The individual chamber shape is elongated in the direction of coiling and partially divided into two lobes by the curved vertical structure of the hookwall, which is an exterior wall that outlines the pseudoumbilicus. The final chamber of the specimen is broken away, leaving the scars of the broken walls which outline the chamber position (Plate 5, figs. 3, 4, 7; Fig. 5).
Only one aperture is present, opening into the elongated trough of the pseudoumbilicus. The aperture is essentially umbilical in position, with the opening facing backwards towards the penultimate chamber. It is bounded by the hook like end of the hookwall at the side closest to the periphery; and on the other extremity, the aperture is bounded by the end of the frontal wall of the final chamber. This wall turns outwards slightly to give the impression of a lip. (This aperture corresponds with Loeblich and Tappan's, 1964, “supplementary aperture” and the “protoforamen” of Hofker, 1952.) The height of the aperture in this specimen is unknown due to the chamber being broken away.
The floor of the apertural opening has seven relatively small tubercules on it. In front of the aperture, a low ridge is capped by 11 slightly larger, pointed tubercules that are tilted towards the centre of the umbilicus. (The possible genesis of this low ridge is discussed later.) On the top of the secondary roof plate of the previous chamber, near to the final apertural opening, another area has 11–12 relatively large tubercules developed (Plate 5, figs. 5, 7).
Older apertures are completely closed off from the exterior by the addition of a large, arched roof plate. This apparently imperforate plate projects further over the pseudoumbilicus than the apertural opening. It attaches to the floor of the pseudoumbilicus inside and adjacent to the hookwall, then arches up and over the hook on that wall, and attaches to the final chamber above the aperture. The roof plate thus covers not only the apertural opening but also the trough of the pseudoumbilicus. In the penultimate chamber, only a shallow curved indentation remains to mark the position of the vertical hookwall. In older chambers this indentation is progressively filled by secondary lamination. In the light microscope, the outline of the roof plates is visible in three to four chambers as a star-like pattern of chamberlets filling the umbilicus around three-quarters of the specimen. In the ESEM™, these roof plates are only faintly visible (Plate 5, fig. 4). However, in spite of the secondary lamination, USNM 383567 does not possess an umbilical boss or plug, an essential characteristic of Discorbis dreheri according to Helmut Bartenstein (personal communication, 30 April 2001).
The intercameral foramen is on the peripheral (outer) side of the curved, vertical hookwall, at the base of the septum with the penultimate chamber. It is a highly ovoid structure formed by secondary resorption (Plate 5, figs. 4, 6, 7). It has been subsequently sealed off by smooth shell material, resembling inner lamella in appearance, so that the protoplasm in the final chamber of this specimen was no longer in contact with the earlier chambers of the specimen at the time of death. The intercameral foramen corresponds to Hofker's (1952) distal “deuteroforamen”, which he considered to be formed by an arching of the suture.
The “aperture”, described as a “low interiomarginal arch” by Loeblich and Tappan (1964) and illustrated in their fig. 637: 2b (reproduced as Plate 5, fig. 2b herein), is a combination of the intercameral foramen described above and a relatively deep depression at the end of the trough (Plate 5, fig. 7). In the light microscope, these two very small structures appear to be one. After the cleaning of the specimen, it was seen in the ESEM™ that two separate depressions are present, separated by the umbilical hookwall (Plate 5, fig. 7). The internal depression within the chamber is the (now-sealed) intercameral foramen (Plate 5, fig. 6), and the external depression is the trough. In USNM 383567, with its broken final chamber walls, the height of the true umbilical aperture is unknown, but the location and width of the aperture can be clearly seen.
Remarks. The usual description of one aperture as running part way along the recessed area behind (= backwards in the direction of coiling) the final chamber is shown to be correct. However, this is the only aperture present; there is no second sutural aperture. The aperture opens into an elongate extension of the pseudoumbilicus, here termed the trough. The aperture is thus an umbilical aperture but faces backwards, towards the earlier chambers, rather than towards the centre of the axis of coiling.
The structure of the chambers on the umbilical side of this specimen was previously completely obscured by a thick layer of gum tragacanth that hindered a correct understanding of the morphology. It can now be seen that the large final chamber, which is broken off, occupies the area interpreted as the fifth-last and sixth-last chambers, plus a part of the area shown as the final chamber, in the 1964 drawing of the specimen (Loeblich and Tappan, 1964, fig. 637: 2b–2c; also in Loeblich and Tappan, 1987, pl. 476, figs. 14–15; reproduced as Plate 5, figs. 2b and 2c herein). The roof plates (“cover-plates” of Hofker, 1952) are imperforate. The term “cover plate” is not used because this is a very different structure in the Rotaliina.
The vertical hookwall has been variously referred to as the “internal partition” (Copestake and Johnson, 2014), “hooklike intracameral plate” (Loeblich and Tappan, 1987), or “toothplate” (Hofker, 1952). Hofker (1952) appears to have been misled by the absence of the final chamber in his specimens. However, as this “partition” is always an external wall in the final chamber, it cannot be correctly referred to as a partition. The use of the term “toothplate” has been extensively discussed by Revets (1993), who presents a case for regarding the toothplate as always an internal structure within the chamber (amongst other characteristics). This is certainly not the case for the “toothplate”/hookwall in Reinholdella, where this structure only secondarily becomes internal after the formation of the next chamber and the associated roof plate which closes off the preceding chamber.
4.4 Specimens from the type sample (Hambühren WA 2, 342 m, Germany)
Material examined. The virtual topotype specimens examined from borehole Hambühren WA 2, 342 m, opalinum zone (Aalenian) were picked by Bartenstein, and later the same sample was repicked and despatched to the author courtesy of Heinz Malz. The picked specimens were mounted on a slide in gum tragacanth; however, the specimens dissolved in the post on the way to the author in Australia. A second group of specimens was sent loose and unmounted in another slide, and many of these unfortunately arrived crushed under the coverslip and broken. Of those that survived, most are very small and variable in preservation and morphology.
Remarks. The specimens comprise at least two species and possibly more. Photographs of these specimens were taken by the author and sent to Bartenstein in March 2001, with specific questions as to which (particularly the biconvex specimen of Plate 6, figs. 1–3) corresponded to Discorbis dreheri Bartenstein (1937). In a letter of 30 April 2001 to the author, Bartenstein indicated that the umbilical knob was a defining characteristic for D. dreheri. He did not indicate if any of those photographs of his picked specimens actually fitted his concept of D. dreheri. Only one specimen supplied from the type sample actually had any indication of a filled umbilicus or a structure that could doubtfully be interpreted as an umbilical knob (M76-07 (SMF XXVII 14538) Plate 6, figs. 1–3). The other specimens belong to forms with open, flat, or deep open umbilici (Plate 6, figs. 4–6, 8–13; Plate 7, figs. 1–11; Plate 8, figs. 1–12) that evidently belong to different species than the specimens displaying a filled umbilicus.

Plate 6(1–13) Discorbis dreheri Bartenstein, 1937. Specimens from the type material (Hambühren WA 2, 342 m, opalinum zone) received from Malz and Bartenstein. SEM photos. (1–3, 7) biconvex keeled form, specimen M76-07 (SMF XXVII 14538). (1) Slightly oblique umbilical view showing last chamber entirely broken away. (2) Edge view (not apertural view), seen from the broken margin of second-last chamber. (3) Spiral view, showing prominent raised thickened intercameral sutures, depressed spiral suture, and peripheral keel. Scale bar 100 µm for (1)–(3). (7) detail of spiral chamber surface of third-last chamber, showing finely perforate chamber surface and thickened raised imperforate intercameral sutures. Scale bars each 10 µm. (4–6) Broken specimen M76B-14 (SMF XXVII 14545). (4) Spiral view showing raised thickened spiral and intercameral sutures and keel. (5) Oblique edge view showing domed spiral surface and thick perforate wall underlying the sutures. (6) Detail of broken spiral chamber wall at lower centre of (5); note the numerous fine perforations. Upper part of photo shows a thin area of etched wall. Scale bar 10 µm. (8) Specimen M74-03 (SMF XXVII 14527) with raised spiral sutures and flat to concave umbilical side; edge (non-apertural) view. (9–11) Specimen M76-09 (SMF XXVII 14540). (9) Oblique edge view, showing the narrowly rounded periphery and flat to slightly convex umbilical side. Spiral side is gently convex. (10) Umbilical view. (11) Spiral view, showing the raised thickened spiral and intercameral sutures. Scale bar 100 µm for (8)–(11). (12–13) Specimen M76B-13 (SMF XXVII 14544), broken (half) specimen. (12) Edge view showing profile of depressed spiral suture, slightly inflated chambers on spiral side, and acute periphery. (13) Broken half of umbilical surface is almost flat. Scale bar for (12)–(13) is 100 µm.
The present author considers that Discorbis dreheri Bartenstein, 1937 cannot be redefined because none of the specimens supplied correspond to the author's original description of the species (Bartenstein, in Bartenstein and Brand, 1937) and because the designated paratypes clearly belong to at least two separate species. It is here regarded as a species nomen dubium.
Description. The morphotypes present comprise the following:
-
Firstly, there is an acute-margined, biconvex species with a domed, largely infilled umbilical area, which merges indistinctly into the chamber surface (Plate 6, figs. 1–2, 5). This morphotype has raised sutures on the spiral side and a very acute margin that in places appears to be almost a blunt keel. The spiral chamber surface is perforate (Plate 6, figs. 6–7). This morphotype is very rare in the collection of some 30 surviving specimens sent to me. It appears to contain some of the features described for Discorbis dreheri in that it has a closed umbilicus with a raised central thickening. However, this area seems to be formed from contiguous or slightly overlapping roof plates and is not divided off from the chamber surface as a separate umbilical plug. It also has raised septal sutures throughout the last whorl on the spiral side and a peripheral keel, whereas in D. dreheri the sutures are described as ridges in the middle of the spiral side. (No keel is mentioned in the description of D. dreheri, and the shape of the periphery is not specified.) The specimen M76-07 (SMF XXVII 14538) (Plate 6, figs. 1–3, 7) has well-developed hookwalls and roof plates in the same pattern as the type specimen of the type species of Reinholdella. This specimen thus seems to belong within the genus Reinholdella, but its placement within the species dreheri Bartenstein is very doubtful due to its biconvex shape, keel, and very prominent sutures.
-
Forms exist which appear transitional between the form above and a more concavo-convex form described below. Three examples of biconvex to almost plano-convex forms are shown (M74-03 (SMF XXVII 14527): Plate 6, fig. 8; M76-09 (SMF XXVII 14540): Plate 6, figs. 9–11; M76B-13 (SMF XXVII 14544): Plate 6, figs. 12–13). These specimens do not have an umbilical plug, but the umbilicus appears to be variably closed by the inward extension of the chambers or in the case of M76-09 (SMF XXVII 14540) (Plate 6, figs. 9–10) by roof plates. The presence of a hookwall could not be clearly established; thus, even the generic placement of these forms is uncertain.
-
The above forms merge into another variant in the Hambühren specimens picked by Bartenstein. This is a narrowly round-margined, convex-concave form with an umbilical hollow and no umbilical plug. This form has flush sutures on the spiral side. This is the most common form with nine photographed specimens in the assemblage supplied from the type sample at Hambühren. It corresponds to the morphology of Loeblich and Tappan's neotype (USNM 383567). However, it does not correspond to the description of the species given by Bartenstein in 1937, in that it has no umbilical plug or “button-shaped thickening” in the middle of the umbilical side. Helmut Bartenstein (personal communication, 2001) reconfirmed this key feature of the description as follows: “In 1937 Discorbis dreheri was introduced as a new species for which the umbilical knob is a characteristic feature distinctive for further determinations” (emphasis Bartenstein's). The current author interprets therefore that this form with a well-rounded margin and open umbilicus lacking a plug does not correspond to Discorbis dreheri. It appears to correspond to Discorbis paraspis (Schwager) (of Bartenstein and Brand, 1937, p. 193, plate 5, fig. 77), which may be referable to Ceratolamarckina. However, the species was illustrated in a small drawing only of the spiral side. The margin of this species was described as well rounded, which corresponds with the specimens illustrated here (Plate 7, figs. 1–3, 5–6; Plate 8, figs. 1–2).

Plate 7(1–11) Discorbis dreheri Bartenstein, 1937. Specimens from the type material (Hambühren WA 2, 342 m, opalinum zone) received from Malz and Bartenstein. SEM photos. (1–4, 8) Concavo-convex form, specimen M76-04 (SMF XXVII 14535). (1) Umbilical view showing open umbilicus. (2) Edge view (non-apertural) with final chamber broken off. Note narrowly rounded periphery and very low trochospire. (3) Oblique edge view with mould of final chamber intact at upper left, before specimen broke during cleaning. Umbilicus is gently concave. (4) Spiral view of gently convex side with partly etched wall. Sutures vary from flush to slightly depressed in the last preserved chamber. (8) Detail of wall surface of last surviving chamber (top right in 4) to show areas of smoother surface with very shallow pitting (centre) and etched areas (right) that expose pores below the surface of the wall. Scale bars on photo for (8) are each 10 µm. (5, 9) Specimen M74-04 (SMF XXVII 14528). (5) Edge view of badly broken specimen to show low trochospiral test and chamber fillings (possibly defined by a thin organic inner lining). (9) Umbilical view to show slightly concave umbilical area and absence of plug. (6–7) Very low trochospiral specimen M74-05 (SMF XXVII 14529). (6) Edge view with narrowly rounded periphery; note the intact peripheral final chamber face, which lacks any apertural openings. (7) Umbilical view; the position of the backward-directed umbilical aperture is visible. (10–11) Specimen M74-01 (SMF XXVII 14525). (10) Umbilical view with final chamber broken away; deep umbilicus and, on two chambers, very prominent roof plates with tubercules. Scale bar 100 µm for (1)–(4), (5)–(7), and (9)–(11). (11) Detailed view of the pseudoumbilical trough (partly filled with sediment) and the roof plate on the second-last chamber, which closes the previous aperture.
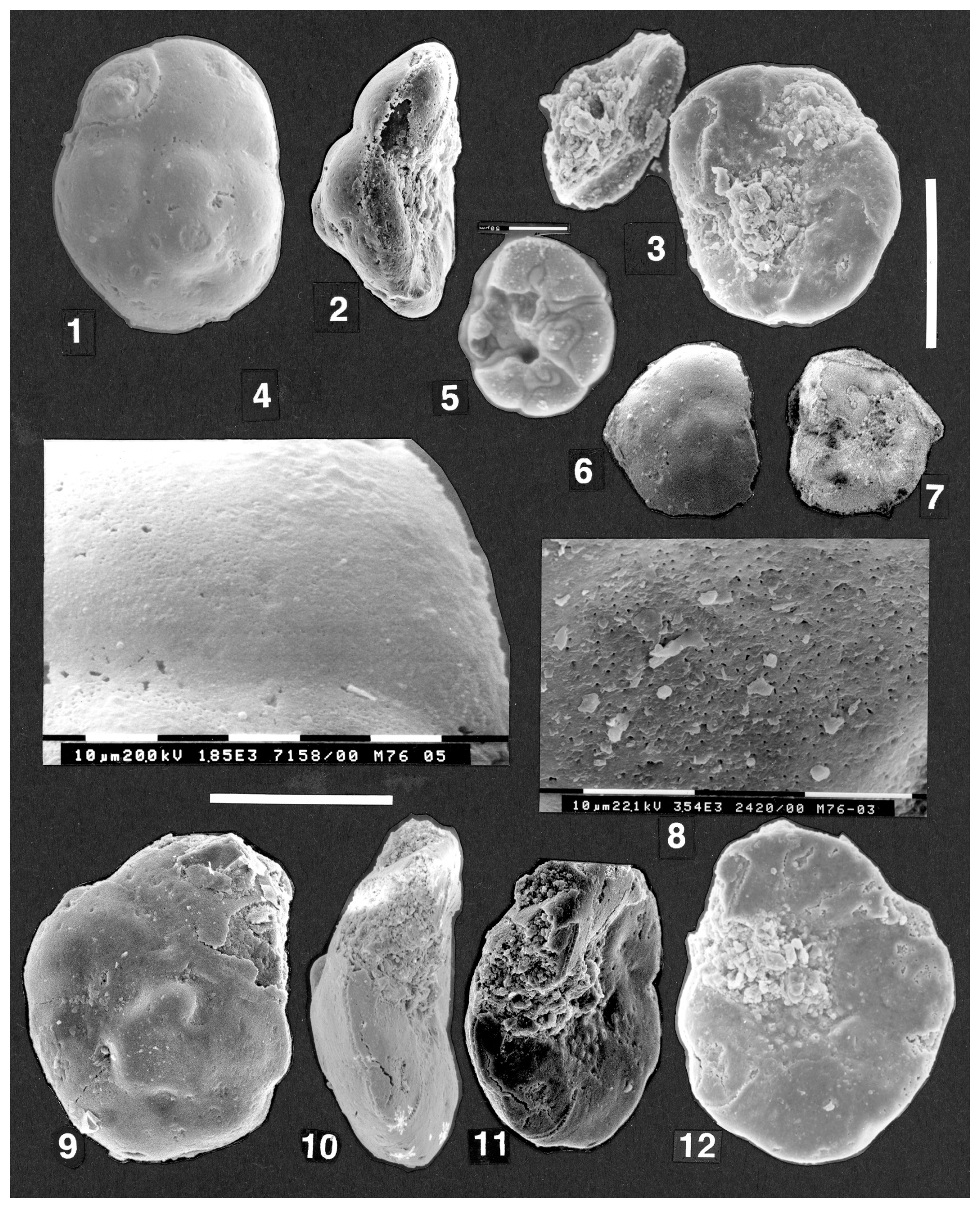
Plate 8(1–4) Discorbis dreheri Bartenstein, 1937. Specimen M76-05 (SMF XXVII 14536); SEM and ESEM™ photos. (1) Spiral view, ESEM™. (2) Edge (non-apertural) view showing the slightly concave umbilical surface and lack of an umbilical plug. (3) Umbilical view. Part of the detached final chamber, broken while attempting to clean the specimen, lies at upper left. This chamber is not shown in (1)–(2). Scale bar to right of (3), 100 µm, for (1)–(3) and (6)–(7). (4) Spiral chamber wall of last preserved chamber seen in (1). Note slight variation in texture from almost smooth areas, slightly lumpy areas, and areas of etching where the very fine underlying pores are exposed. (5) Organic lining within chambers and roof plates from a specimen in which the wall has largely dissolved whilst in transit. The specimen subsequently disintegrated. ESEM™ photo, scale bar 50 µm. (6–7) Discorbis dreheri Bartenstein, 1937. Specimen M76-11 (SMF XXVII 14542). (6) Spiral view of small specimen, ESEM™. (7) Umbilical view, surface badly etched to partly dissolved. Hookwall and area of roof plate clearly visible on chamber filling of last preserved chamber, ESEM™. (8–12) Discorbis dreheri Bartenstein, 1937. Specimen M76-03 (SMF XXVII14534), SEM photos. (8) Spiral surface of earliest chamber (at centre of 9) showing fine perforations reaching surface; the number of these is possibly increased by etching of the surface. Scale bars are each 10 µm. (9) Spiral view showing gently inflated chambers and depressed spiral and intercameral sutures. (10) Edge view to show narrowly rounded periphery and slightly concave umbilical surface. (11) Oblique umbilical and edge view showing concave, open umbilicus, with pustules penetrated by pores. Broken final chamber wall is clearly visible at bottom. Top of specimen cut off in SEM photograph. (12) Umbilical view. Scale bar 100 µm for (9)–(12).
How broadly the genus Reinholdella should be defined is unclear at present, as only two specimens from the type material of the type species have been available for this study. The issue of whether the umbilical dome seen on the holotype of the type species (F.410-1) continuously varies on other specimens to become a partially separated umbilical dome, as on specimen F.410-2, is critical to deciding what other species should be included within the genus. A study of additional material from the type area is therefore required, but enquiries indicated that none could be located.
Clearly the concept of Reinholdella should not include forms with an open umbilicus. Brotzen (1948) described Reinholdella as “distinguished… from Lamarckina by the lack of an umbilicus.”
Also critical to the definition of Reinholdella is the number of external apertures in the last chamber. Hofker (1952) claimed that two apertures were present, one (his protoforamen) on the umbilical side in a loop-shaped indentation of the final suture and one (his deuteroforamen) “at the ventral suture of the last formed chamber near to the margin.” However no such second aperture is present in the holotype, which has an unbroken wall. The presence of this second aperture is shown only in drawings by Hofker (1952), which appear to be interpretations. In discussions between Heinz Malz and Helmut Bartenstein, Bartenstein said that he had been helping Hofker with his research on the “toothplate apertures” (Hofker, 1951) but had resigned from this discussion after he criticised Hofker's drawings, which did not correspond with the specimens under observation. Hofker had then stated that if the last chamber and the aperture were well preserved, they would look in the way he had drawn them (Heinz Malz, personal communication, 23 September 1999). It appears that Hofker was working on specimens with broken or poorly preserved final chambers (as is the case with most Reinholdella) and that he misinterpreted the internal foramen as an external aperture.
A complete re-assessment of the approximately 30 species at present included in Reinholdella is beyond the scope of this study. However, some brief remarks and illustrations are given on a stratigraphically important form to illustrate the umbilical and apertural problems outlined above.
-
Reinholdella macfadyeni (ten Dam, 1947)
Plate 9, figs. 1–11.
-
1947 Asterigerina macfadyeni ten Dam: 396–397, fig. 1a–c.
Material examined. About 20 specimens from Rudd's Pit, Grantham, Lincolnshire, England (“Upper Liassic, ?bifrons zone” (Toarcian) according to the label), courtesy of the British Museum of Natural History. The sample was collected on 25 November 1938 by William A. Macfadyen and was evidently part of the sample sent to ten Dam. An assemblage slide in BMNH drawer T105, picked by Charles G. Adams in 1958, is marked “Topotype material”. Although ten Dam indicated that his new species was common in this locality, he did not designate it as the type locality. The figured specimens are stored under the catalogue numbers NHMUK PM PF 75498–75504.
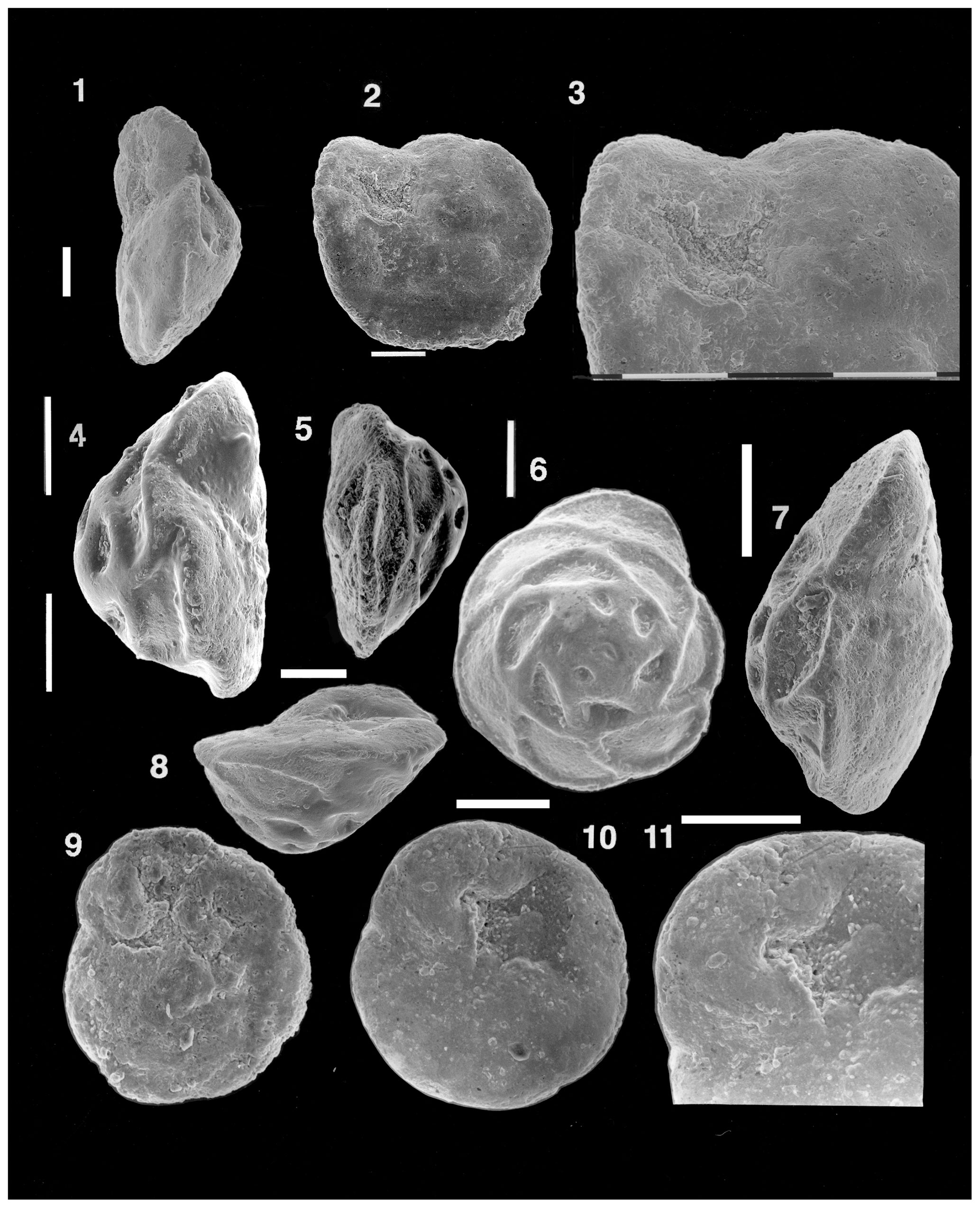
Plate 9(1–11) Reinholdella macfadyeni (ten Dam, 1947). (1–3) Specimen M43-09, NHMUK PM PF 75499. (1) Slightly oblique edge view showing the final suture without an apertural opening. (2) Umbilical view of same specimen with a preserved but etched final chamber, a low umbilical dome, and a single interiomarginal umbilical aperture. (3) Detailed view of the final chamber, tilted slightly towards the viewer, showing no visible apertural opening on the final suture. (4) Specimen M43-12, NHMUK PM PF 75502: edge view to show the plano-convex test and the absence of any aperture on the intact final chamber suture. (5–6) Specimen M43-11, NHMUK PM PF 75501. (5) Edge view, slightly oblique, showing the low umbilical dome and nearly plano-convex test. (6) Spiral view to show the prominent raised and thickened sutures. (7) Specimen M43-10, NHMUK PM PF 75500. (7) Edge view to show biconvex test shape and low umbilical dome. Well-preserved final chamber shows no apertural opening on its suture near the periphery. (9) Specimen M43-8, NHMUK PM PF 75498. (9) Slightly etched specimen, umbilical view, showing the small but intact final chamber at top left, the small roof plate over the second-last chamber, and the low umbilical dome separated from the last four chambers by a shallow groove. (8, 10–11) Specimen M43-14, NHMUK PM PF 75504. (8) Edge view looking towards the indentation of the interiomarginal aperture. (10) Umbilical view to show absence of a central umbilical dome. Prominent lip over the aperture is visible, adjacent to pseudoumbilical area with tubercules. Roof plate on the second-last chamber almost reaches centre of specimen. (11) Detail of intact final chamber. No apertural opening is visible on the final suture adjacent to the margin. SEM photos. Scale bars of 100 µm for figs. (1)–(11).
(An assemblage slide from the type locality was also examined by the author in the British Museum of Natural History. The specimens of Reinholdella macfadyeni in this slide were more poorly preserved in the vicinity of the apertural complex, and none of these specimens were borrowed for photography.)
Remarks. Reinholdella macfadyeni is an important species in the British and northern European Jurassic, ranging from the Upper Pliensbachian to Lower Aalenian (Copestake and Johnson, 2014). Specimens examined for this study are uniform as to the features of the spiral side: 2.5 to 3 whorls of chambers, with strongly raised, thickened spiral and intercameral sutures, the latter obliquely directed backwards. The sutural thickening on early chambers almost covers the chamber surfaces in the first whorl. In edge profile, specimens display variability in shape, ranging from almost plano-convex to (less commonly) unequally biconvex, with the spiral side being the more convex. Most variability is seen in the features on the umbilical side. A few specimens lack an umbilical dome (M43-14; NHMUK PM PF 75504: Plate 9, figs. 10–11), having an open pseudoumbilical area behind the last chamber. Some specimens have a smooth umbilical dome above the axis of coiling (specimen M43-10, NHMUK PM PF 75500: Plate 9, fig. 7), without separation from the chamber surface of the chambers, except for the final one. Other specimens have a low umbilical dome that is clearly separated (by a groove) from both the roof plates and the chamber surface of the last, penultimate, and third-last chambers (M43-11; NHMUK PM PF 75501: Plate 9, fig. 5; M43-8: NHMUK PM PF 75498: Plate 9, fig. 9). While some specimens have the umbilical features seen on the type specimen of Reinholdella (F.410-1), others resemble ten Dam and Reinhold's second specimen (F.410-2) in having a partially developed umbilical plug. The best-preserved specimens display no second aperture, which according to Hofker (1952) should be present close to the periphery on the final suture of the last chamber (M43-10, NHMUK PM PF 75500: Plate 9, fig. 7; M43-14, NHMUK PM PF 75504: Plate 9, figs. 8, 10–11). It is noteworthy that none of the eight species of Reinholdella described and figured by Ohm (1967) have two apertures.
-
The diagnosis of the genus Reinholdella is herein described and illustrated based on the type specimen of the genotype, Reinholdella dreheri Brotzen, 1948, which is the specimen F.410-1, identified and illustrated as Asterigerina dreheri (Bartenstein, 1937) by ten Dam and Reinhold (1942). The type specimen is biconvex, with an umbilical side that rises to a low dome above the axis of coiling. There is a very shallow pseudoumbilical area on the slope of this dome and no separate umbilical plug. There is only one aperture, a pseudoumbilical opening facing obliquely backwards, towards the second-last chamber. There is no second aperture (that should be marked by a break) on the final chamber wall near the margin. There is no roof plate (= “cover plate”, “supplementary chamberlet”) over the final chamber. Consequently, the structure here termed the hookwall (= “internal partition”; “inner partition” of Brotzen, 1948; “toothplate” of Hofker, 1951, 1952) is shown to be always an external wall of the most recently formed chamber. Only later, after the formation of the next chamber, does the hookwall become an internal structure. In the penultimate chamber, part of the frontal wall on the umbilical side of the test has been resorbed prior to formation of the next chamber. The position of the resorbed wall is marked by a low ridge, often capped by tubercules (Plate 1, fig. 8; Plate 2, figs. 2–3; Plate 4, figs. 1–2; Plate 5, fig. 7; Figs. 2, 3, 5). The hookwall of the next chamber is deposited over the top of this low ridge, crossing the ridge at an oblique angle. This implies resorption of the entire previous frontal wall in some cases and the formation of a new septum that is concave in the direction of chamber growth. The intercameral foramen is a resorbed oval opening in this new septum. Resorption is mentioned in passing in the literature on aragonitic foraminifera (for example, by McGowran, 1966), but its importance in chamber construction appears to have been somewhat overlooked. In all specimens photographed for this study, the roof plates (“cover plates” of other authors) are imperforate, with only irregular corrosion pits from diagenesis.
-
The holotype of Reinholdella dreheri Brotzen, 1948 (F.410-1) does not possess an umbilical plug in the sense of a separate raised structure. Instead, the low dome in the centre of this side of the test is continuous with the slope of chambers and roof plates and seems to be formed by secondary lamination at the same time as the formation of successive roof plates over earlier chambers.
-
The second specimen in the type slide (F.410-2) is a different species, with a moderately separated umbilical plug and differences in test shape and chamber features. Further study of type material, if any can be found, is highly desirable to establish whether continuous variation exists between the two species.
-
The question of whether the type species of Reinholdella (F.410-1) is conspecific with Discorbis dreheri Bartenstein, 1937 does not affect the status of specimen F.410-1 as the type specimen of the type species of Reinholdella.
From the evidence presently available, it is concluded that the two Dutch specimens are not conspecific. Under Articles 11.10, 69.2.4, and 70.3 of the Code of Zoological Nomenclature, the type species of Reinholdella is Reinholdella dreheri Brotzen, 1948.
-
How broadly the genus Reinholdella should be defined is unclear at present, as only two specimens from the type material of the type species have been available for this study. The holotype provides a central anchor point for the genus, but the issue of whether the umbilical dome seen on the holotype of the type species (F.410-1) continuously varies on other specimens to become a partially separated umbilical dome, as on specimen F.410-2, is critical to deciding what other species should be included within the genus. A study of additional material from the type area is therefore required, but enquiries indicated that none could be located.
-
In light of the diagnosis of Reinholdella given here, species of “Reinholdella” with a rounded periphery and an open, unfilled umbilicus (e.g. R. brandi, Hofker, 1952) will need to be reassigned to another genus. Species with umbilical plugs as separate structures on the exterior of the test (e.g. some specimens of R. macfadyeni (ten Dam, 1947)) are provisionally accepted as belonging within Reinholdella. However further study is needed on whether a distinction should be made between umbilical plugs and domed, “continuously filled” umbilical areas. This requires the study of additional specimens from ten Dam and Reinhold's material, if any still exists.
No data sets were used in this article.
The supplement related to this article is available online at https://doi.org/10.5194/jm-44-169-2025-supplement.
The author has declared that there are no competing interests.
Publisher’s note: Copernicus Publications remains neutral with regard to jurisdictional claims made in the text, published maps, institutional affiliations, or any other geographical representation in this paper. While Copernicus Publications makes every effort to include appropriate place names, the final responsibility lies with the authors.
The present investigation would not have been possible without the taxonomic and ESEM™ expertise of Stefan Revets, then at the University of Western Australia, Department of Geology and Geophysics, who generously agreed to borrow and photograph the type slide of Reinholdella dreheri and the neotype of Discorbis dreheri on behalf of the author. Stefan Revets also provided taxonomic guidance, encouragement, and frequent stimulating discussions on foraminiferal morphology. However, the final opinions expressed herein are the responsibility of the author. The curators of the Geologische Stichting Haarlem, and of the Smithsonian Institution, Washington, D.C., are thanked for the loans of the specimens of Asterigerina dreheri (F.410) and Discorbis dreheri (USNM 383567) for photography. The curators of the Natural History Museum, London, are thanked for the loan of specimens of Reinholdella macfadyeni from the collection.
Helmut Bartenstein kindly picked additional specimens of Discorbis dreheri from the type material from Hambühren, which is stored in the Senckenberg Museum, and made these available for study through Heinz Malz (Bramsche, Germany). Heinz Malz and Helmut Bartenstein also corresponded with the author on the characteristics of the species discussed here, and this correspondence is attached in the Supplement. David Haig of the University of Western Australia provided encouragement, facilities, and the freedom to pursue the topic. The Centre for Microscopy and Microanalysis at the University of Western Australia is thanked for providing access to electron microscope facilities. Philip Copestake twice reviewed different versions of the manuscript, thereby greatly improving the quality and conclusions of this study. Stefan Revets and an anonymous reviewer commented extensively on the final version, ensuring adherence to the International Code of Zoological Nomenclature, and they are particularly thanked for this.
This paper was edited by Laia Alegret and reviewed by Stefan Revets and one anonymous referee.
Bartenstein, H. and Brand, E.: Mikro-paläontologische Untersuchungen Stratigraphie des nordwest-deutschen Lias und Doggers: Abhandlungen der Senckenbergischen Naturforschenden Gesellschaft, Abhandlung 439, 1–224, 1937.
Brotzen, F.: The Swedish Paleocene and its foraminiferal fauna, Årsbok Sveriges Geologiska Undersökning, 42, 1–140, 1948.
Brotzen, F.: Correction of genotype, The Micropaleontologist, 3, p. 30, 1949.
Copestake, P. and Johnson, B.: The Hettangian to Toarcian, in: Stratigraphical Atlas of Fossil Foraminifera, edited by: Jenkins, D. G. and Murray, J. W., 2nd Edn., 129–188, ISBN 0-85312-210-5, 1989.
Copestake, P. and Johnson, B.: Lower Jurassic Foraminifera from the Llanbedr (Mochras Farm) Borehole, North Wales, UK, Monograph of the Palaeontographical Society, London, 1–403, pls 1–21 (Publ. 641, part of Vol. 167 for 2013), 2014.
Gradstein, F. M.: Biostratigraphy and biogeography of Jurassic Grand Banks foraminifera, in: First International Symposium on Benthonic Foraminifera of Continental Margins, Part B, Paleoecology and Biostratigraphy, edited by: Schafer, C. T. and Pelletier, B. R., Maritime Sediments, Special Publication No. 1. Halifax, Canada, 790 pp., 557–583, 1976.
Hofker, J.: The Toothplate Foraminifera, Archives Néerlandaises de Zoologie, 8, 353–372, 1951.
Hofker, J.: The Jurassic Genus Reinholdella Brotzen (1948) (Foram.), Paläont. Z., 26, 15–29, 1952.
Hottinger, L.: Illustrated glossary of terms used in foraminiferal research, Carnets Géol., Memoir 2006/02 (CG2006-M02), 43 pp., 2006.
ICZN (International Code of Zoological Nomenclature): The International Trust for Zoological Nomenclature, 4th Edn., London, UK, 306 pp., ISBN 0 85301 006 4, 1999.
Loeblich Jr., A. R. and Tappan, H.: Treatise on Invertebrate Paleontology, Part C, Protista 2, Sarcodina, Geol. Soc. Am., 2, C1–900, 1964.
Loeblich, A. R. Jr. and Tappan, H.: Foraminiferal Genera and their Classification, Van Nostrand Reinhold, New York, 2 vols, 1–970 + 1–212, 847 pl., 1987.
McGowran, B.: Bilamellar walls and septal flaps in the Robertinacea, Micropaleontology, 12, 477–488, 1966.
Ohm, U.: Zur Kenntnis der Gattungen Reinholdella, Garantella und Epistomina (Foramin.), Palaeontogr. Abt. A, 127, 103–188, 1967.
Revets, S. A.: The Foraminiferal Toothplate, A Review, J. Micropalaeontol., 12, 155–169, https://doi.org/10.1144/jm.12.2.155, 1993.
ten Dam, A.: A new species of Asterigerina from the Upper Liassic of England, J. Paleontol., 21, 396–397, 1947.
ten Dam, A. and Reinhold, T., Some foraminifera from the lower Liassic and the lower Oolitic of the Eastern Netherlands, Geol. Mijnbouw, 4, 8–11, 1942.
- Abstract
- Introduction
- Historical background and nature of the problem
- Description of the type specimen of the type species of the genus Reinholdella
- The nature of Discorbis dreheri Bartenstein
- Remarks on concepts of Reinholdella
- Conclusions
- Data availability
- Competing interests
- Disclaimer
- Acknowledgements
- Review statement
- References
- Supplement
- Abstract
- Introduction
- Historical background and nature of the problem
- Description of the type specimen of the type species of the genus Reinholdella
- The nature of Discorbis dreheri Bartenstein
- Remarks on concepts of Reinholdella
- Conclusions
- Data availability
- Competing interests
- Disclaimer
- Acknowledgements
- Review statement
- References
- Supplement