the Creative Commons Attribution 4.0 License.
the Creative Commons Attribution 4.0 License.
 | 22 Sep 2025
| 22 Sep 2025
Detailed conodont data from the Olenekian–Anisian boundary interval of the GSSP candidate section at Deşli Caira, Romania
Martyn L. Golding
The Olenekian–Anisian boundary (OAB) interval is an important time in Earth's history, reflecting the last phase of marine ecosystem recovery in the aftermath of the end-Permian mass extinction. Despite this, the Global Boundary Stratotype Section and Point (GSSP) for the base of the Anisian remains undefined. The first appearance of the conodont Chiosella timorensis has been proposed as a potential index for the boundary; however, the discovery of this conodont with ammonoids traditionally considered to be Spathian (late Olenekian) has generated doubts about its suitability as a proxy. At the GSSP candidate section at Deşli Caira, Romania, the OAB was previously drawn at the base of bed GR7, which contained the first occurrence of Ch. timorensis; however, additional collecting has shown that Spathian ammonoids persist higher in the section, and recent studies have instead placed the OAB at the base of bed 822A, approximately 3 m above the previous placement. The conodont faunas from this higher interval are less well studied than those from the lower level; furthermore, the beds in this part of the section have now been subdivided in more detail than they were previously, with bed 822 alone now being split into six sub-beds. Existing conodont faunas are only recorded as coming from bed 822, so their position relative to the new subdivision is not certain. In order to improve the precision of conodont correlation around the potential higher position for the OAB, 10 new samples were collected from bed 821 to bed 824. In addition to characterizing the fauna throughout the OAB interval, these new collections also enable the evolution of late Spathian and early Anisian conodont species to be understood in more detail.
- Article
(4000 KB) - Full-text XML
- Comment
- BibTeX
- EndNote




The Olenekian–Anisian boundary (OAB) is currently the focus of a concerted international effort to define the base of the Anisian stage with a Global Boundary Stratotype Section and Point (GSSP). Recently, numerous studies have appeared on the sedimentology, palaeontology, geochemistry, magnetostratigraphy, and geochronology of several OAB sections, including those at Kçira in Albania (Muttoni et al., 2019), Wantou in China (Chen et al., 2020), and especially Deşli Caira in Romania (Grădinaru and Gaetani, 2019; Golding, 2021; Grădinaru, 2022; Horacek and Grădinaru, 2023; Balini et al., 2024). The need to define this boundary has become particularly pressing with the recognition that the Anisian represents the last phase of recovery of marine organisms from the end-Permian mass extinction, along with the re-establishment of normal oceanographic and climatic conditions and of complex marine ecosystems (e.g. Hofmann et al., 2011; Sun et al., 2012; Chen and Benton, 2012; Wignall, 2015; Song et al., 2015; Liang et al., 2016; Pietsch et al., 2019; Friesenbichler et al., 2021a, 2021b; Echevarría and Ros-Franch, 2024); therefore, the present lack of certainty in the correlation of this boundary hampers our ability to understand the rates and mechanisms of this biotic recovery.
The Anisian stage was first recognized based on limestone belonging to the Gutensteinerkalk at Großreifling in Austria (Waagen and Diener, 1895; Arthaber, 1896; Assereto, 1971; Summesberger and Wagner, 1972). The scope of the stage was subsequently expanded and re-defined by Arthaber (1906) and Pia (1930), with the original Anisian of Waagen and Diener (1895) being regarded as the Upper Anisian and the beds below being included in the Lower Anisian (encompassing the Hydaspian of Waagen and Diener, 1895; see schemes of Pia, 1930; Rosenberg, 1959; and Zapfe, 1974, 1983). As the type region lacked ammonoids in the lower part of the Anisian, the basal part of the stage was later further refined in Greece and Türkiye, where ammonoids were more abundant (Assereto, 1974). The sections in the Marathovouno hillock area on the island of Chios in Greece and on the Kocaeli Peninsula in Türkiye became the stratotypes for the Aegean and Bithynian sub-stages, respectively. Additional studies at Chios on the ammonoid and conodont faunas, along with chemostratigraphy and magnetostratigraphy, have been published (Bender, 1970; Nicora, 1977; Assereto et al., 1980; Fantini Sestini, 1981; Gaetani et al., 1992; Jacobshagen et al., 1993; Muttoni and Rettori, 1994; Muttoni et al., 1994, 1995, 1996; Mertmann and Jacobshagen, 2003), and the sections on this island remain important for understanding the OAB; however, they are no longer considered to be suitable candidates for the GSSP due to condensation and mixing of ammonoid faunas, the absence of terminal Spathian ammonoid faunas, and faulting in the vicinity of the boundary (Bucher, 1989; Muttoni et al., 1996).
The base of the Aegean sub-stage (and, consequently, the base of the Anisian stage) was initially recognized on the basis of the first occurrence of the ammonoid genus Paracrochordiceras (Assereto, 1974). Other ammonoid genera have been used to recognize the boundary in different regions, including Aegiceras, Japonites, and Paradanubites (Gaetani, 1993; Balini et al., 2010; Ogg et al., 2014); however, more recent studies at Deşli Caira advocating a boundary based on ammonoid faunal turnover have yet to define a marker species (Balini et al., 2024). Other potential markers that have been proposed to aid recognition of the OAB include a peak in the δ13C values (Grădinaru et al., 2007; Horacek and Grădinaru, 2023) and the shift from reversed-polarity to normal-polarity magnetozones (the base of magnetozone MT1n; Hounslow et al., 2007; Hounslow and Muttoni, 2010).
The first appearance of the conodont Chiosella timorensis has been proposed as a suitable biotic proxy for the boundary by several authors (Nicora, 1977; Assereto et al., 1980; Orchard and Tozer, 1997; Orchard et al., 2007a, b; Orchard, 2010; Muttoni et al., 2019; Tong et al., 2019; Chen et al., 2020), and this species is recognized in the OAB interval at all major sections of this age around the world. However, Ch. timorensis has also in many instances been found with ammonoids traditionally considered to be Olenekian (Nakazawa and Bando, 1968; Mirăuţă, 1974, 2000; Atudorei et al., 1997; Konstantinov and Klets, 2009; Goudemand et al., 2012; Grădinaru, 2022), and its taxonomy, morphological variation, and stratigraphic range have been poorly understood, leading to uncertainty about its relationship with the morphologically similar Ch. gondolelloides; the first occurrence of this species is normally recorded either from the same beds as Ch. timorensis (e.g. Chen et al., 2020) or from slightly below (e.g. Orchard et al., 2007a, b). Recent morphometric analysis of specimens from Deşli Caira and elsewhere has indicated that Ch. gondolelloides is most likely the juvenile form of Ch. timorensis (Wu et al., 2025), an interpretation that pushes the first appearance of Ch. timorensis lower into what is currently regarded as the Olenekian.
The only current, formally proposed candidate section for the GSSP for the base of the Anisian is at Deşli Caira in Romania. This section has been studied extensively since the early 20th century (Kittl, 1908; Simionescu, 1910, 1911), with substantial work subsequently published on the fossil ammonoids and nautiloids (Grădinaru, 2000, 2022; Grădinaru and Sobolev, 2006; Balini et al., 2023, 2024), brachiopods (Grădinaru and Gaetani, 2019), and microfossils (Mirăuţă, 1974, 2000; Mirăuţă et al., 1993; Crasquin-Soleau and Grădinaru, 1996; Grădinaru and Ivanova, 2002; Grădinaru et al., 2006; Orchard et al., 2007a; Sebe et al., 2013; Forel and Grădinaru, 2021; Golding, 2021) and on the chemostratigraphy (Atudorei, 1999; Grădinaru et al., 2007; Horacek and Grădinaru, 2023) and magnetostratigraphy (Grădinaru et al., 2007) of the section. In 2006, Grădinaru et al. formally proposed using the first occurrence of Ch. timorensis as the proxy to define the base of the Anisian in the section. In addition to Deşli Caira, research is ongoing at two other possible candidate sections, Wantou in southern China (Wang et al., 2005; Ovtcharova et al., 2006, 2015; Galfetti et al., 2007, 2008; Ogg, 2019; Chen et al., 2020) and Kçira in Albania (Muttoni et al., 1996; Germani, 1997; Meço, 1999, 2010; Muttoni et al., 2019); however, neither of these sections have yet been formally proposed as candidate sections.
Subsequent to the initial definition of the boundary in the Deşli Caira section at the first occurrence of Ch. timorensis in sample GR7 (Orchard et al., 2007a), Grădinaru and Gaetani (2019) reported Olenekian ammonoids from above this level in the section and suggested that the boundary should be revised in light of these reports (although these ammonoids were not illustrated or described). These authors recommended that the boundary should be moved from below sample GR7 to the base of bed 822A, approximately 3 m higher. Grădinaru and Gaetani (2019) also introduced a new numbering scheme for the section, with beds 821 and 822 divided into several new sub-beds; some of these beds and sub-beds were subsequently subdivided again, with the most recent numbering scheme being utilized by Balini et al. (2024). This new scheme allows much greater precision in determining the sequence of biotic events in this part of the section but also makes it difficult to tell which bed the archival samples collected prior to the new numbering should be assigned to.
The current paper presents the results of new conodont sampling from the stratigraphically higher proposed OAB interval of the Deşli Caira section, utilizing the new bed numbering scheme. The new sampling has allowed the conodont fauna around this modified OAB level to be assessed and accurately tied to the ammonoid faunas recovered and described in detail by Balini et al. (2024). Ranges of several conodont species have been extended due to new occurrences, and the suitability of the revised, stratigraphically higher level for marking the OAB has been assessed, as has the utility of the first occurrence of Ch. timorensis to serve as a biotic proxy for the boundary.
The studied section is located on the Deşli Caira hill in southeastern Romania, near the coast of the Black Sea (Fig. 1), with the base of the section at 45°04′27′′ N, 28°48′08′′ E (Grădinaru et al., 2007). The Deşli Caira section is approximately 60 m thick, with the lowest 20–25 m occurring on the southeastern slope of the hill (Section A of Grădinaru et al., 2007). These rocks are entirely Spathian in age and consist of dolostone overlain by Hallstatt-type limestones (Grădinaru et al., 2007). The samples discussed in the present paper come from the next-youngest unit, preserved on the southwestern slope of the hill (Section B of Grădinaru et al., 2007), which is composed of nodular limestone grading up into Hallstatt-type limestone, thought to have been deposited on a deep submerged plateau (Atudorei et al., 1997; Forel and Grădinaru, 2021). These rocks are commonly considered to be part of the Agighiol Limestone (Atudorei et al., 1997; Grădinaru, 2005; Grădinaru et al., 2007), and this terminology is frequently used in publications on the Deşli Caira section (e.g. Grădinaru et al., 2007; Grădinaru and Gaetani, 2019; Forel and Grădinaru, 2021; Golding, 2021; Horacek and Grădinaru, 2023; Balini et al., 2024); however, other authors have preferred to use the term nodular and bioturbated limestone for these rocks, although this terminology is less widespread in the literature (Baltres, 2005; Baltres et al., 2020). A fault has been recognized in section B, with its trace estimated in Figs. 4 and 5 of Balini et al. (2024), although its position between beds 822A2 and 822B is uncertain. The rocks of the Deşli Caira section were formed as part of a carbonate platform developed on the central eastern part of the Tulcea Unit, a major tectonostratigraphic unit of the North Dobrogean Orogen, which was formed by subduction of the Palaeo-Tethys Ocean prior to being thrust over Eurasia during the Cimmerian Orogeny (Grădinaru, 1995, 2000).
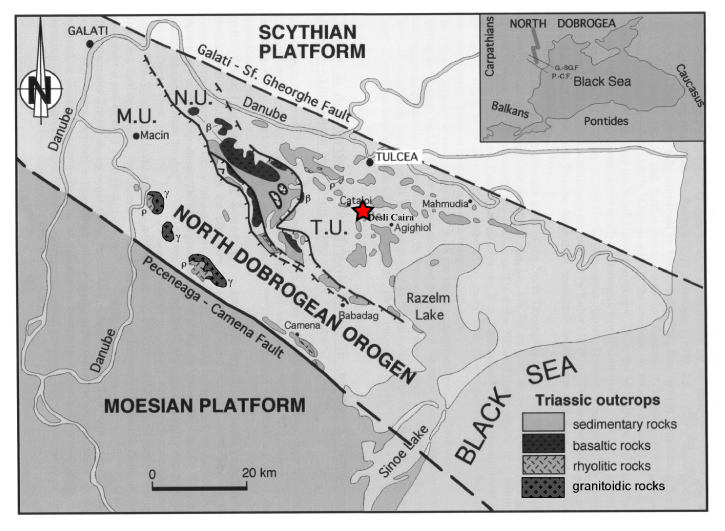
Figure 1Locality map showing the location of the Deşli Caira Section (marked by a red star) in the North Dobrogean Orogen of eastern Romania; the location with respect to the Black Sea is shown in the inset (modified from Grădinaru et al., 2007).
During the summer of 2022, 10 new conodont samples were collected from the higher OAB interval identified by Grădinaru and Gaetani (2019). These samples covered a total of less than 3 m of stratigraphic height and encompassed all of the beds and sub-beds identified by Grădinaru and Gaetani (2019) between beds 821 and 824 inclusive (Fig. 2). The stratigraphic position of the samples can be tied precisely to the 8xx numbering system for previous samples collected from the section (Grădinaru et al., 2007; Orchard et al., 2007a). However, their position relative to other numbering schemes (e.g. 2xx and 9xxx) of these papers is less certain, with only relative position known. The samples were each approximately 2 kg in weight and consisted of walnut-sized chunks of red Hallstatt-type limestone. Sample numbering follows the format 22-GWA-DC-xxx, where -xxx refers to the bed or sub-bed that the sample was collected from.
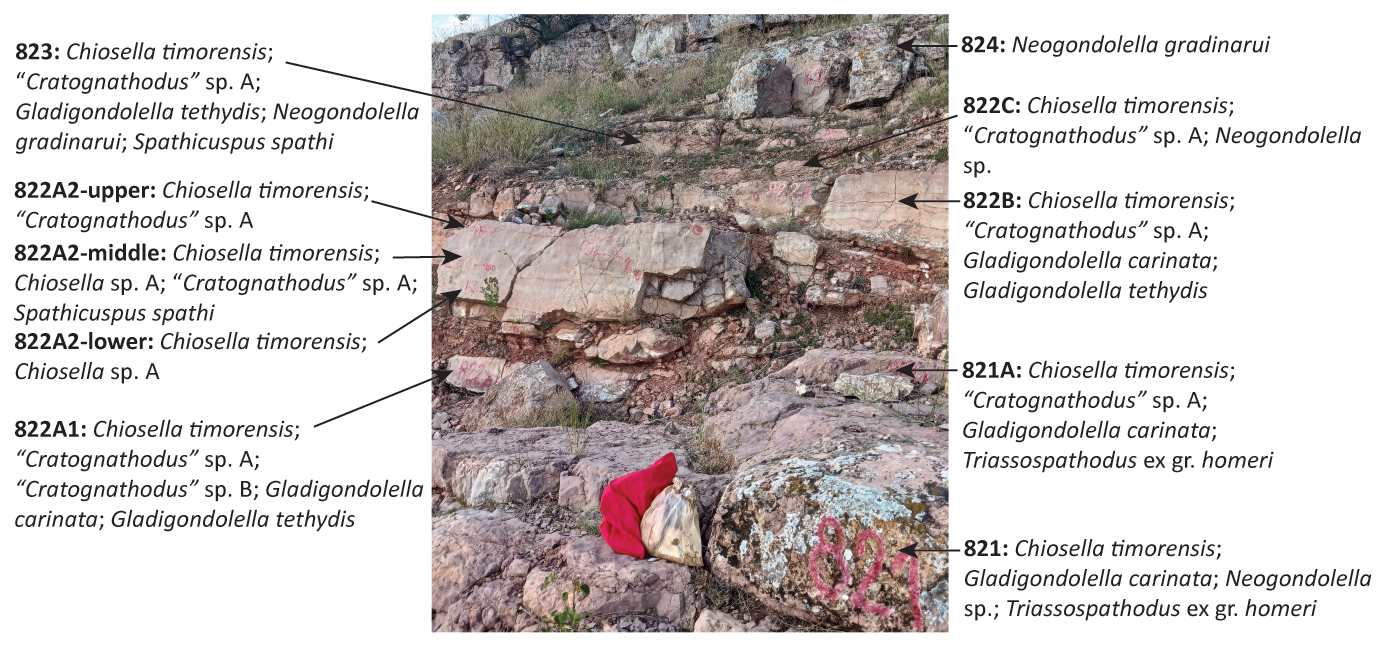
Figure 2Outcrop photo of the Deşli Caira section (Section B of Grădinaru et al., 2007), illustrating the new bed numbering system, and the stratigraphic levels of samples collected from the section (location of each sample within a bed is approximate). The species found within each sample are listed alongside. Note that Balini et al. (2024) indicate the presence of a fault within bed 822B, which appears to be a crack within the bed to the left of the arrow labelling this bed. See sample bag on bed 821 (approximately 30 cm) for scale.
Samples were delivered to the conodont laboratories of the Geological Survey of Canada in Calgary and Vancouver, where they were processed using standard techniques as outlined in Stone (1987). Firstly, samples were acidized in buffered dilute acetic acid for 10–14 d, before the dissolved residue was passed through sieves to capture the 90–850 µm fraction. This isolated fraction was then rinsed, dried, and put through heavy liquid separation using lithium metatungstate with a specific gravity of 2.85 kg L−1. The heavy fraction was picked by hand underneath a light microscope, and conodonts were photographed using a Hitachi TM3000 scanning electron microscope housed at the Geological Survey of Canada in Vancouver. All figured specimens are reposited in the National Type Collections of the Geological Survey of Canada in Ottawa.
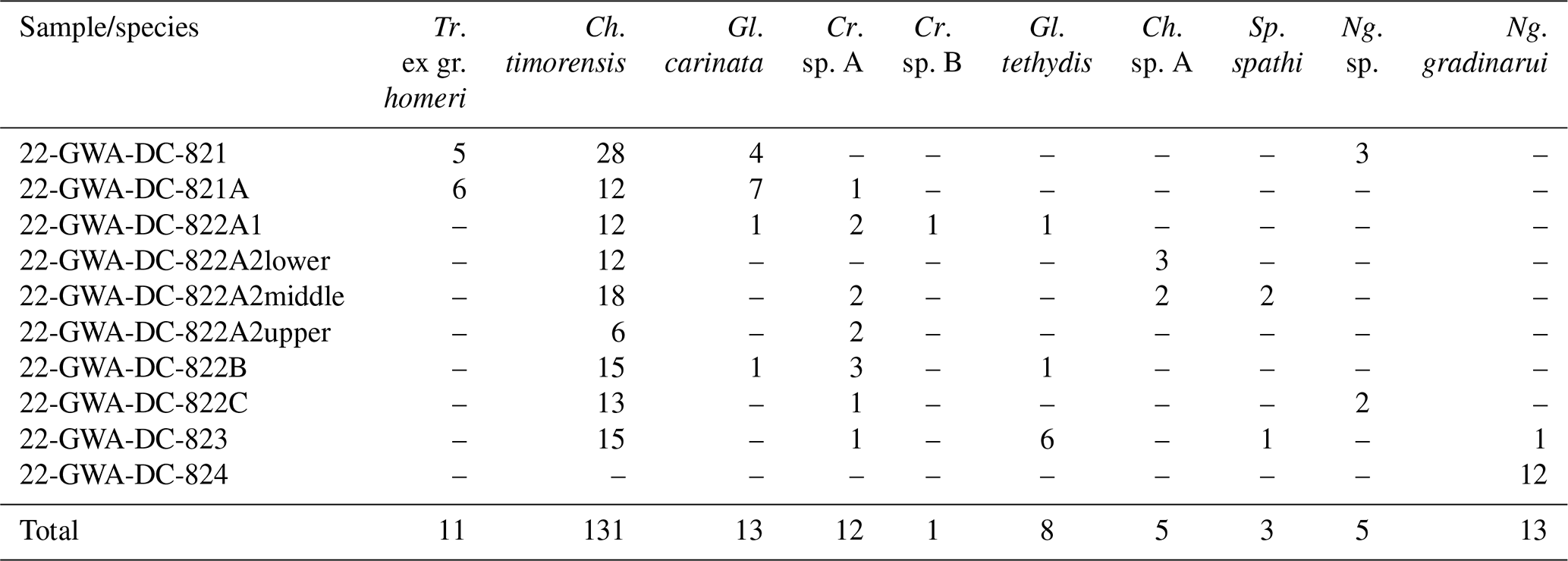
All 10 of the new conodont samples collected as part of this study were productive, with more than 200 conodont P1 elements being recovered. The specimens belonged to 10 different species, and a list of the species found in each sample is presented in Table 1; their stratigraphic positions are also illustrated in Figs. 2 and 3.
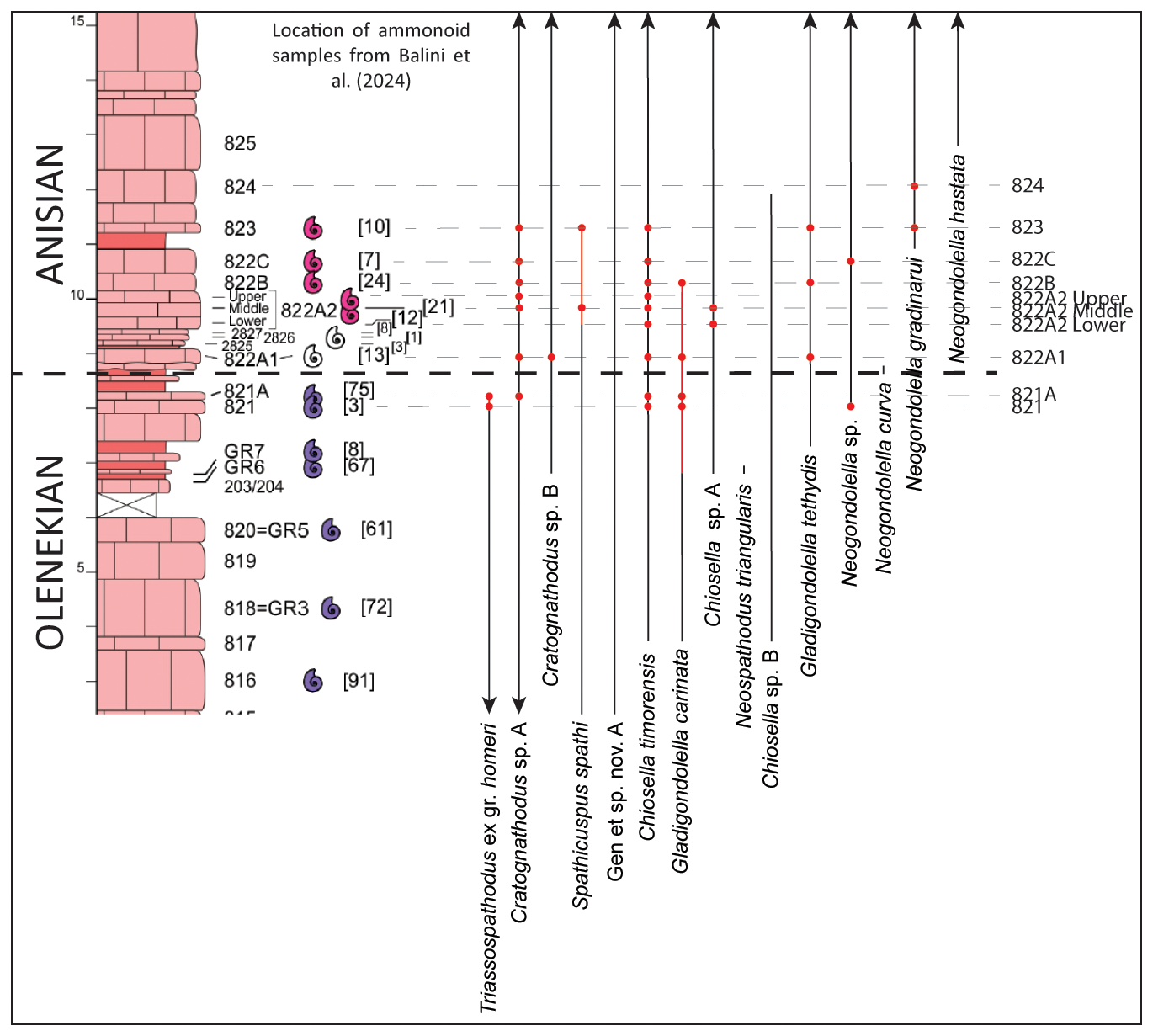
Figure 3Stratigraphic range chart for the conodont species recovered from the new sampling at Deşli Caira. The stratigraphic height of new samples is indicated by red dots, and range extensions of species are indicated with red lines. Existing conodont ranges are based on data in Orchard et al. (2007a) and Golding (2021) and are shown in black. Arrows indicate a continuation of the species range in the section beyond the part illustrated here. The position of the Olenekian–Anisian boundary is tentative and is placed between beds 821 and 822A to reflect the opinion of Grădinaru and Gaetani (2019) and to mark the base of the Olenekian–Anisian ammonoid faunal turnover interval of Balini et al. (2024). The lithologic log, position of ammonoid samples, and number of ammonoid specimens (in square brackets) are all based on Balini et al. (2024).
Table 1Conodont specimen counts by sample. Tr.: Triassospathodus; Ch.: Chiosella; Gl.: Gladigondolella; Cr.: Cratognathodus; Sp.: Spathicuspus; Ng.: Neogondolella.
-
22-GWA-DC-821. This fauna is dominated by specimens of Triassospathodus ex gr. homeri (Fig. 4). The species Tr. homeri was first described from the Spathian of the former GSSP candidate section at the Marathovouno hillock area in Greece by Bender (1970). It has subsequently been described from Spathian and Anisian strata from around the world, including Pakistan (Sweet, 1970; Matsuda, 1985), India (Chhabra and Sahni, 1981; Matsuda, 1983), Primor'ye (Zakharov and Rybalka, 1987), the Pamirs (Dagys, 1990), southern China (Duan, 1987), Oman (Orchard, 1995), and North America (Orchard and Tozer, 1997; Henderson et al., 2018). However, there are also several allied species of Triassospathodus described from the Spathian which can be difficult to differentiate from Tr. homeri, including Tr. symmetricus, Tr. brochus, and T. sosioensis (Orchard, 1995). Together, these species are often grouped as Tr. ex gr. homeri, and that practice is followed here. Larger collections would be necessary to differentiate these species more confidently. In addition to Tr. ex gr. homeri, this sample contains specimens of Gladigondolella carinata (Fig. 4), Chiosella timorensis, and the earliest examples of Neogondolella sp. to have been reported from the Deşli Caira section. The specimens of Ch. timorensis recovered from this sample include examples of what has previously been called Ch. gondolelloides (Orchard et al., 2007a); however, the analyses of Goudemand et al. (2012) and Wu et al. (2025) have demonstrated that Ch. gondolelloides is mostly synonymous with Ch. timorensis, representing smaller specimens of that species. Therefore, all specimens with the morphology of either Ch. gondolelloides or Ch. timorensis have been referred to the latter species in the present study. Gl. carinata was also originally named for specimens from Chios (Bender, 1970; Gaetani et al., 1992) and has subsequently been described from OAB sections in Romania (Orchard et al., 2007a) and China (Orchard et al., 2007b; Lehrmann et al., 2015; Chen et al., 2020), although its relationship with other species of Gladigondolella is uncertain due to differences in unpublished multielement collections from the published reconstructions of Gladigondolella tethydis (Orchard, 2005; Ishida and Hirsch, 2011; Kılıç et al., 2013; Golding and Kılıç, 2024).
-
22-GWA-DC-821A. The fauna from this sample is similar to that of 22-GWA-DC-821, consisting primarily of Ch. timorensis (Fig. 4), Gl. carinata, and the highest representatives of Tr. ex gr. homeri (Fig. 4) in the section. These species are joined by examples of Cratognathodus sp. A. This species was first named by Orchard at al. (2007a, b) based on material from Deşli Caira and Guandao. Subsequently, based on a statistical analysis of the co-occurrence of elements, Golding and Kılıç (2024) demonstrated that, rather than being a separate genus, Cratognathodus actually belongs to the multielement apparatus of Gladigondolella, as previously suggested by Ishida and Hirsch (2011) and Kılıç et al. (2013). Despite this, the morphological variability in the cratognathodid element appears to have stratigraphic utility during the OAB interval, with Cr. sp. B occurring stratigraphically higher in the sections at Deşli Caira and Guandao than Cr. sp. A (Orchard et al., 2007a, b). Therefore, in the present paper, the specific names of Cratognathodus elements are retained.
-
22-GWA-DC-822A1. In addition to Gl. carinata, Ch. timorensis (Fig. 4), and Cr. sp. A (Fig. 4), as found in lower parts of the section, this sample also contains specimens of Cr. sp. B (Fig. 4) and Gl. tethydis (Fig. 4). Both of these species have lower occurrences at Deşli Caira. This record of the species significantly extends its range at Deşli Caira from previous reports. The first occurrence of Gl. tethydis has previously been recognized as an important datum in the vicinity of the OAB at several sections, including Deşli Caira (Orchard et al., 2007a), Guandao (Orchard et al., 2007b; Goudemand et al., 2012), and Kçira (Muttoni et al., 2019). However, the relative occurrence of this species is variable; in most sections it occurs slightly higher than Ch. timorensis, but at Kçira it occurs below Ch. timorensis (Meço, 1999; Muttoni et al., 2019), together with late Spathian ammonoids (Grădinaru, 2022), and the species has recently been reported from strata as old as early Spathian in Oman (Leu et al., 2023).
-
22-GWA-DC-822A2lower. A relatively depauperate sample, the only species recovered from this level were Ch. timorensis and Ch. sp. A (Fig. 4). The latter species was named by Orchard et al. (2007a, b) based on material from Deşli Caira and Guandao and has subsequently been recognized from other sections, including Mingtang, Wantou, and Youping (Chen et al., 2020; Shunling Wu, personal communication, 2024). Utilizing this new material, work is underway to fully describe this species and its relationship to other species and morphotypes of Chiosella.
-
22-GWA-DC-822A2middle. This sample contains specimens of Ch. timorensis, Ch. sp. A, Cr. sp. A, and Spathicuspus spathi (Fig. 4); this is the lowest of the new collections to contain the latter species; however, representatives of this species have previously been found lower in the section as well (Orchard et al., 2007a). Sp. spathi was first described from Pakistan (Sweet, 1970) and subsequently reported from India (Garzanti et al., 1995), Albania (Meço, 1999; Muttoni et al., 2019), North America (Orchard, 2005, 2022), China (Orchard, 2005; Orchard et al., 2007b; Brühwiler et al., 2009; Chen et al., 2013, 2015, 2023; Lehrmann, et al., 2015; Liang et al., 2016; Wu et al., 2024), Oman (Agematsu et al., 2008; Chen et al., 2019), and Japan (Maekawa et al., 2021), where it is most commonly recorded in the Spathian, but occasional reports of Anisian occurrences also exist (e.g. Meço, 1999).
-
22-GWA-DC-822A2upper. This is another depauperate sample, containing only a few specimens of Ch. timorensis and Cr. sp. A.
-
22-GWA-DC-822B. This sample contains specimens of Ch. timorensis, Cr. sp. A, Gl. carinata, and Gl. tethydis. A fault in the section with approximately 80 cm of offset may mean that this sample is actually from bed 822A2 (Marco Balini, personal communication, 2024; see Balini et al., 2024, Fig. 5 for illustration of this fault); therefore, a new sample has been collected from bed 822B to confirm the location of sample 22-GWA-DC-822B, and the results from this new sample will be reported on in the future. The fauna of sample 22-GWA-DC-822B is very similar to those from samples both above and below; however, it currently possesses the last specimens of Gl. carinata found in the section, so the uppermost limit of this species may have to be revised down from bed 822B to bed 822A2 if the new samples from bed 822B cannot corroborate this occurrence (see Fig. 3).
-
22-GWA-DC-822C. This sample contains specimens of Ch. timorensis (Fig. 4), Cr. sp. A, and Neogondolella sp.
-
22-GWA-DC-823. As well as specimens of Gl. tethydis (Fig. 4), Ch. timorensis, and Cr. sp. A, this sample contains the youngest specimens of Sp. spathi found at the Deşli Caira section, extending the range of this species significantly upwards (more than 3 m) compared with previous reports. This sample also contains the first examples of Ng. gradinarui recovered in the new samples (Fig. 4), although older specimens of this species have previously been reported from this section (Orchard et al., 2007a; Golding, 2021). Abundant specimens from this sample and the overlying 22-GWA-DC-824 allow the recognition that examples of Ng. cf. constricta reported in Golding (2021) are actually juvenile examples of Ng. gradinarui. This removes uncertainty about the origins and stratigraphic range of Ng. constricta, confirming that the younger species first appears in the Pelsonian (as suggested by Orchard and Golding, 2024) and not the Bithynian. In addition to its occurrence in Romania, Ng. gradinarui has now also been recognized from Anisian strata in sections at Guandao (Golding, 2021) and Datuguan (Chen et al., 2023) in southern China.
-
22-GWA-DC-824. As mentioned above, this sample is dominated by specimens of Ng. gradinarui (Fig. 4). This is the stratigraphically highest of the new samples collected, and, although this is the first major influx of Neogondolella species in the Deşli Caira section, Chiosella returns to once again dominate higher samples in the section (see Discussion).
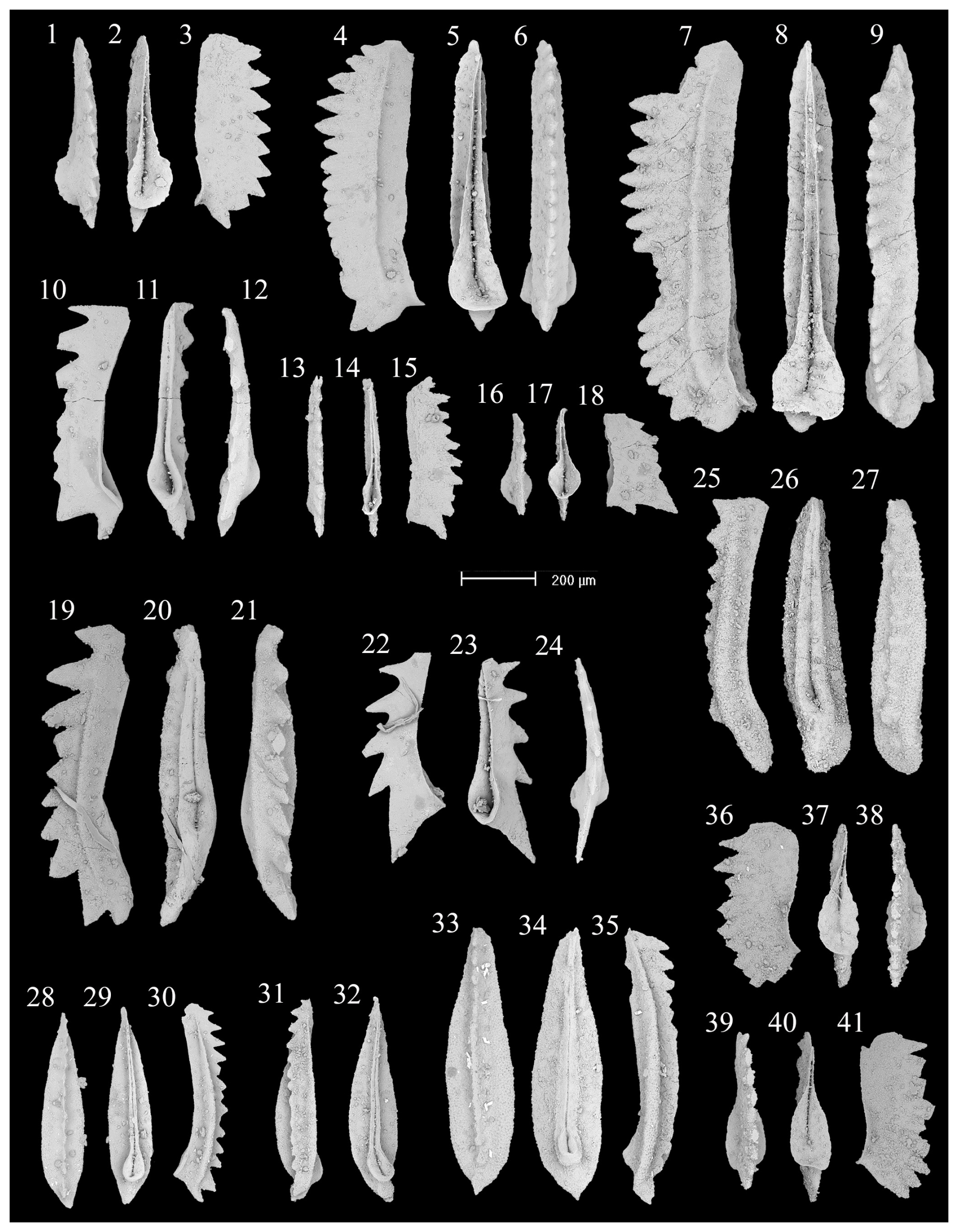
Figure 4SEM photos of a selection of conodonts recovered from the new sampling at the Deşli Caira section. (1–9) Chiosella timorensis (Nogami): (1)–(3) GSC Cur. No. 139713, from sample 22-GWA-821A, (4)–(6) GSC Cur. No. 139714, from sample 22-GWA-822A1, (7)–(9) GSC Cur. No. 139715, from sample 22-GWA-822C. (10–12) Cratognathodus sp. A (sensu Orchard et al., 2007a), GSC Cur. No. 139716, from sample 22-GWA-822A1. (13–15) Chiosella sp. A (sensu Orchard et al., 2007a), GSC Cur. No. 139717, from sample 22-GWA-822A2lower. (16–18) Spathicuspus spathi (Sweet), GSC Cur. No. 139718, from sample 22-GWA-822A2middle. (19–21) Gladigondolella carinata Bender, GSC Cur. No. 139719, from sample 22-GWA-821. (22–24) Cratognathodus sp. B (sensu Orchard et al., 2007a), GSC Cur. No. 139720, from sample 22-GWA-822A1. (25–27) Gladigondolella tethydis Müller, GSC Cur. No. 139721, from sample 22-GWA-823. (28–35) Neogondolella gradinarui Golding and Orchard: (28)–(30) GSC Cur. No. 139722 and (33)–(35) GSC Cur. No. 139723, both from sample 22-GWA-824, (31) and (32) GSC Cur. No. 139724, from sample 22-GWA-823. (36–41) Triassospathodus ex gr. homeri (Bender): (36)–(38) GSC Cur. No. 139725, from sample 22-GWA-821A, (39)–(41) GSC Cur. No. 139726, from sample 22-GWA-821. All photos x80 magnification.
The new conodont collections from Deşli Caira allow refinement of the stratigraphic range and taxonomic scope of several species. The stratigraphic ranges of Triassospathodus ex gr. homeri, Spathicuspus spathi, and Gladigondolella carinata have all been extended upwards compared with previous reports, whereas the stratigraphic range of Neogondolella sp. has been extended downwards (see Fig. 3). Integration of new samples with some existing samples remains imprecise, due to uncertainty over which sub-bed certain samples were originally sourced from. Nevertheless, important comments can be made on the composition of the conodont fauna at Deşli Caira and its significance for the recognition of the OAB in this section. The remainder of the Discussion will make reference to both the new and existing samples from this section.
5.1 Faunal changes at the Deşli Caira section
By plotting the relative proportions of conodont genera per sample, it is possible to recognize two major changes in conodont faunal composition which occur at Deşli Caira (Fig. 5). The lowest part of the section is dominated by specimens of Triassospathodus and Cratognathodus until sample 9038 (originally collected by Viorel Atudorei and reported on in Orchard et al., 2007a), where there is a large influx of Chiosella. Sample 9038 is located approximately 4.5 m below the ammonoid-based OAB of Grădinaru and Gaetani (2019), Grădinaru (2022), and Balini et al. (2024). This level represents the first appearance of Chiosella in the section, and this genus remains a significant component of the conodont fauna throughout the rest of the section.
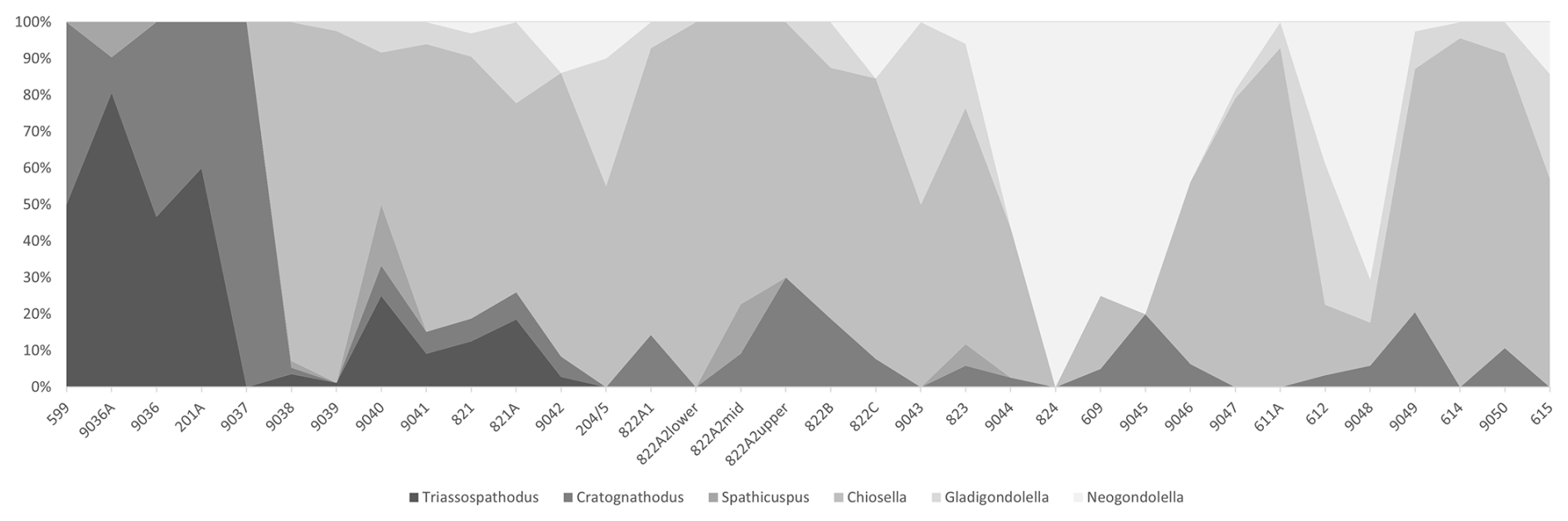
Figure 5Chart illustrating the relative proportions of conodont genera in each sample from the Deşli Caira section, including both new and existing samples (see Orchard et al., 2007a, and Golding, 2021, for the stratigraphic position of existing samples). Percentages were calculated on raw counts of the original specimens and are plotted on the y axis. The sample number is on the x axis, increasing in stratigraphic height to the right. The relative position of the samples is not to scale. Particularly large influxes of Chiosella (in sample 9038) and Neogondolella (in samples 9044 and 824) can be observed.
The second significant change in conodont faunal composition occurs in samples 9044 (collected by Viorel Atudorei) and 824 (this study), where there is a large increase in the relative proportion of Neogondolella. Although this genus first appears in sample 821 (this study), it is a small component of the fauna until samples 9044 and 824, which both occur approximately 2 m above the ammonoid-based OAB. The percentage of Neogondolella in higher samples fluctuates, but it remains a large if not dominant component of the fauna throughout the rest of the section.
These two levels indicate times of significant change in the conodont faunas during the OAB interval at Deşli Caira and suggest that species belonging to either of these two genera would be most suitable for a conodont-defined position for the OAB in this section.
5.2 Taxonomic scope and stratigraphic range of Chiosella timorensis
The taxonomy of Chiosella in the earliest Anisian is a matter of ongoing study. In earlier reports of the conodonts from Deşli Caira, two species of Chiosella were distinguished: the older Ch. gondolelloides and the younger Ch. timorensis (Grădinaru et al., 2006; Orchard et al., 2007a). However, work by Goudemand et al. (2012) on specimens from Nevada demonstrated that both morphologies (gondolelloides: no or short rib; timorensis: longer rib that reaches posterior of the element) were present in the oldest described sample of Chiosella (from the Spathian Haugi Zone) and show a degree of gradation between them that precludes specific separation. Instead, these authors proposed that the name Ch. gondolelloides should be restricted to the holotype and that other specimens previously referred to this species should subsequently be referred to Ch. timorensis. Recent morphological work by Wu et al. (2025) on specimens of Chiosella from several sections around the world supports this conclusion; not only is the length of rib not a significant morphological factor in populations of Chiosella, it is also clear that those specimens with short or no ribs (Ch. gondolelloides morphology) are almost always the smaller specimens in a sample. Larger specimens have longer, more robust ribs; therefore Ch. timorensis and Ch. gondolelloides likely represent the adult and juvenile ontological stages, respectively, of a single species. This observation is particularly true at Deşli Caira, where faunas are dominated by small elements of Chiosella with no rib or a short rib; only larger specimens have ribs that reach the posterior end of the element, and the largest may have ribs that reach the posterior on both sides, sometimes surrounding it (Fig. 4). In the present study, all specimens previously referred to Ch. gondolelloides are included in the scope of Ch. timorensis, as reflected in Table 1 and Fig. 3.
The Chiosella taxonomic story is further complicated by the introduction of new species: Ch. n. sp. A, Ch. n. sp. B (both initially defined in Orchard et al., 2007a, b), and the differentiation at Wantou and Youping sections of Ch. timorensis s.l. and Ch. timorensis s.s. (Chen et al., 2020). Although Ch. n. sp. A and Ch. n. sp. B do appear to be valid species, the question of Ch. timorensis s.s. versus Ch. timorensis s.l. is harder to unravel, as the definition of the latter remains vague. Introduced as a term to incorporate all forms of Ch. timorensis that diverged from the holotype, it includes specimens with arched profiles, rounded rather than triangular basal cavities, and varying fusion of the blade and carina. Many of these morphotypes are present in the collection of Goudemand et al. (2012) from the Haugi Zone, suggesting that all of these may have appeared at the same time and therefore have little stratigraphic utility. At the Wantou section, both Ch. timorensis s.s and Ch. timorensis s.l. first occur in the same sample (Chen et al., 2020). Further work on the faunas of the Wantou section is needed to be able to discriminate between different morphotypes of Ch. timorensis and to determine if there is any stratigraphic separation of morphotypes. At Deşli Caira, it is not possible at present to recognize an OAB defined by the FAD of Ch. timorensis s.s. vs. Ch. timorensis s.l., as the lowest samples in the section that contain Ch. timorensis already show wide morphological variability. Again, further work on these faunas will be necessary.
The occurrence of Ch. timorensis (including specimens which could be referred to Ch. timorensis s.s.) in the Haugi Zone of Nevada (Goudemand et al., 2012) demonstrates that the FAD of this species must be below the Japonites beds in Tethys; this is true at the Deşli Caira section, where the first occurrence of Ch. timorensis in sample 9038 is approximately 4.5 m below the Olenekian–Anisian ammonoid turnover observed by Balini et al. (2024). As mentioned previously, Nakazawa and Bando (1968) were the first to suggest that Ch. timorensis co-occurred with Spathian ammonoids in the collections of Nogami (1968) from Timor, and its suitability as a proxy for the OAB has been questioned by several authors since (e.g. Mirăuţă, 1974, 2000; Atudorei et al., 1997; Konstantinov and Klets, 2009; Goudemand et al., 2012; Grădinaru, 2022). Adoption of the FAD of Ch. timorensis as the proxy for definition of the base of the Anisian would by definition require several ammonoid species presently considered to be confined to the Olenekian to have their ranges extended into the Anisian.
5.3 Taxonomic scope and stratigraphic range of Gladigondolella tethydis
Gladigondolella tethydis has previously been suggested as a possible alternative conodont proxy for the OAB (e.g. Golding, 2021). As demonstrated by Golding and Kılıç (2024), the most recent multielement reconstruction of Gladigondolella suggests that Cratognathodus elements also belong to this genus. Previously at Deşli Caira, these two genera have been treated separately, with the first specimens of Cratognathodus occurring in sample 509 (collected by Eugen Grădinaru) and the first specimens of Gladigondolella occurring in sample 9038 (at the same time as the revised Ch. timorensis), approximately 4 m higher in the section. The recognition that these two genera belong to the same multielement apparatus therefore extends the range of Gladigondolella far below the OAB interval at Deşli Caira and makes the position of the first appearance of Gl. tethydis uncertain, as elements of Cr. sp. A and B may belong to the apparatus of Gl. tethydis or Gl. carinata. This suggests that Gl. tethydis is therefore not a suitable proxy for the OAB, as its first occurrence may well be older than that of Ch. timorensis in the Deşli Caira section. Furthermore, Leu et al. (2023) have recently recognized specimens of Gl. tethydis from the lowest part of the Spathian in Oman, indicating that the first appearance of this species is indeed much older than the OAB and not suitable as a proxy for the boundary.
5.4 Taxonomic scope and stratigraphic range of Neogondolella gradinarui
As mentioned above, the appearance of abundant Neogondolella in sample 9044 represents a real change in the composition of the conodont fauna at Deşli Caira, although Chiosella is still a large component of the fauna at this level and above. Previously, Golding (2021) suggested that a species of Neogondolella may be suitable as an alternative proxy for the OAB and indicated that either Ng. curva or Ng. gradinarui could be potential candidates. Balini et al. (2024) noted that the first occurrence of Ng. curva in the section is very close to the position of the ammonoid-defined OAB interval. However, the new sampling described in this paper has failed to recover additional specimens of this species, and the occurrence of Ng. curva at Deşli Caira is therefore still limited to just one sample. This argues against utilizing Ng. curva as an OAB proxy, as its stratigraphic range is uncertain.
In contrast, Ng. gradinarui has now been recognized in the lowermost Anisian at several sections globally (Deşli Caira, Guandao, Datuguan; Golding, 2021; Chen et al., 2023), and our understanding of its growth stages and stratigraphic range has been greatly improved by the results of the new sampling at Deşli Caira. It now appears that what Golding (2021) previously referred to as Ng. cf. constricta actually represents the juvenile form of Ng. gradinarui, with growth leading to a fusion of the cusp and posterior denticle noted in the smaller forms and a broadening of the posterior platform, which is narrower and more constricted in the juveniles (Fig. 4). At Deşli Caira, Ng. gradinarui first occurs just above (less than 2 m) the level of the OAB proposed on the basis of ammonoids in sample 9043 and would be the most suitable of the Neogondolella species for a proxy of the OAB.
5.5 Position of the Olenekian–Anisian boundary at the Deşli Caira section
At Deşli Caira, the first occurrence of Anisian ammonoids as recognized by Grădinaru and Gaetani (2019), Grădinaru (2022), and Balini et al. (2024) coincides with the base of magnetozone MT3n (Balini et al., 2024; red line in Fig. 6), and the first occurrence of Chiosella timorensis coincides with the beginning of the peak of an increase in carbon isotope values observed by Horaceck and Grădinaru (2023; green line in Fig. 6). In contrast, the first occurrences of both Gladigondolella tethydis and Neogondolella gradinarui do not coincide with clear auxiliary abiotic events at Deşli Caira. It would seem to be appropriate to choose either level (green or red) in defining the OAB at Deşli Caira, as both are supported by multiple proxies and both would allow straightforward correlation with other important sections worldwide.
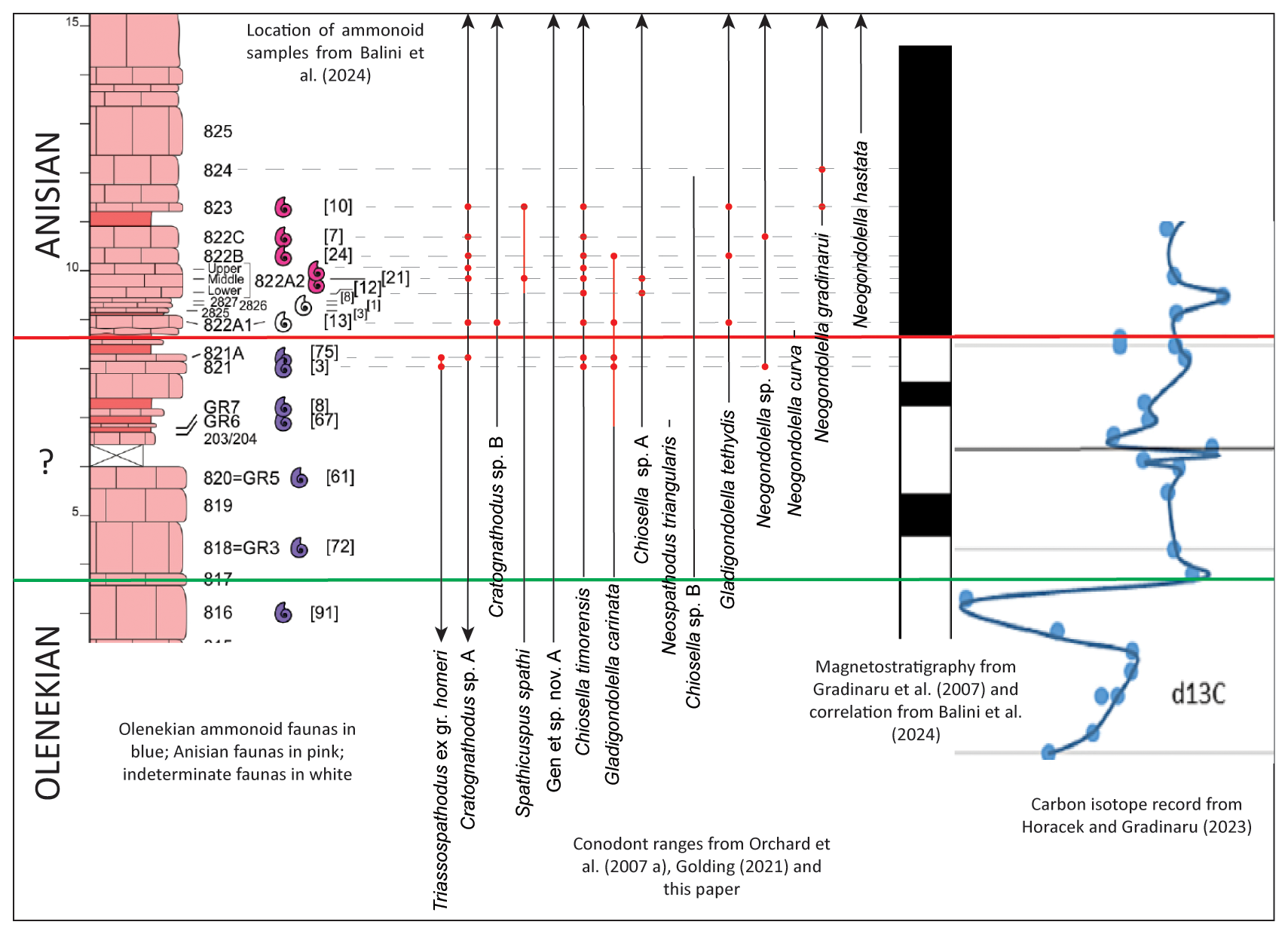
Figure 6Correlation of conodont occurrences from the Deşli Caira section (red dots) with ammonoids, carbon isotope values, and magnetostratigraphy. The two horizontal lines (red and green) reflect two different possibilities for the placement of the OAB: the red line coincides with the shift from Olenekian to Anisian ammonoid faunas as demonstrated by Grădinaru and Gaetani (2019) and Balini et al. (2024) and with the base of the MT3n magnetozone (Hounslow et al., 2007; magnetostratigraphy from Grădinaru et al., 2007). The green line coincides with the first occurrence of Chiosella timorensis in the section, together with the beginning of the peak of a carbon isotope excursion (Horacek and Grădinaru, 2023). The position of the Olenekian–Anisian boundary is adjusted from Fig. 2 to reflect the two different options for placement of the boundary.
In fact, investigation of the available data indicates that the sequence of biotic and abiotic events surrounding the OAB seems to be consistent at most of the major sections encompassing this time interval. At Deşli Caira, Kçira, and Wantou, as well as at Youping, Mingtang, and Guandao, the first occurrence of Ch. timorensis (in the broader sense defined by Goudemand et al., 2012, and Wu et al., 2025, and advocated for in this paper) in each section is at or close to the onset of the peak positive shift in carbon isotopes that can be observed in all of these sections (Horacek and Grădinaru, 2023; Muttoni et al., 2019; Chen et al., 2020; Shunling Wu, personal communication, 2024). At Deşli Caira, both the first occurrence of Ch. timorensis and the beginning of the peak carbon isotope excursion are below the earliest Anisian ammonoid faunas (see position of isotope excursion in Horacek and Grădinaru, 2023, and position of ammonoids in Balini et al., 2024), and this occurrence of Anisian ammonoids is approximately coincident with the base of a normal-polarity magnetozone at this section (MT3n; see correlation in Balini et al., 2024, fig. 15). Equivalent magnetozones (WT2, GU2 and Kc2n) in the Wantou, Guandao and Kçira sections (where Lower Anisian ammonoids are lacking) appear to be in a comparable position with regard to the first occurrence of Ch. timorensis and the carbon isotope excursion in these sections (see Balini et al., 2024, Fig. 15; Chen et al., 2020, Fig. 7; Muttoni et al., 2019, Fig. 8). The sequence of biotic and abiotic events also appears to be in the same order at Chios; however, the faulting near to the boundary makes the recognition of the order of events in this section uncertain (see Balini et al., 2024, Fig. 15, and Chen et al., 2020, Fig. 7).
This consistent succession of events at OAB sections around the world means that the boundary interval can be recognized with some confidence globally. The question merely remains which event should serve as the boundary itself and at what horizon the base of the Anisian should be placed. Since both levels indicated in Fig. 6 are supported by multiple proxies, either would be suitable. However, at present, no single ammonoid species has been proposed as a biotic proxy for the GSSP of the OAB, although such a proposal is reportedly in progress (Balini et al., 2024). Furthermore, many sections with good records of the OAB interval nonetheless lack ammonoids (e.g. Wantou, Guandao). As such, the most promising proxy for the definition of the boundary remains the first occurrence of Ch. timorensis, even though this would require several ammonoid species to be re-classified as ranging from Olenekian to basal Anisian in age and some of the currently recognized ammonoid zones would straddle the boundary.
New samples from the Deşli Caira section in Romania allow greater characterization of the conodont fauna around the OAB interval in this section. These collections cover the revised position of the boundary as recognized by the appearance of Anisian ammonoids and as discussed by Grădinaru and Gaetani (2019), Horacek and Grădinaru (2023), Balini et al. (2024), and others and utilize the new subdivision of beds around this boundary. The new samples demonstrate that there is no significant conodont turnover or speciation event at this level. The most significant conodont events in the section are the influx of Chiosella, marked by the first occurrence of Ch. timorensis (in the broader definition advocated by Goudemand et al., 2012; Golding, 2021; and Wu et al., 2025) much lower in the section than the first Anisian ammonoids, and the influx of Neogondolella, in particular the species Ng. gradinarui (which has now been recognized from several sections around the world and has had its growth stages better illustrated herein), which first occurs much higher in the section than the first Anisian ammonoids. The ranges of several conodont species have been extended in the section, with the last occurrences of Triassospathodus ex gr. homeri, Spathicuspus spathi, and Gladigondolella carinata all revised upwards and the first occurrence of Ng. spp. revised downwards.
The correlation between conodont biostratigraphy, ammonoid biostratigraphy, carbon isotope chemostratigraphy, and magnetostratigraphy suggests that a similar sequence of several biotic and abiotic events can be observed at Deşli Caira as at other OAB sections around the world, including Chios, Kçira, Wantou, and Guandao. This confirms that Deşli Caira would be very suitable as a GSSP for the OAB, as all major biotic and abiotic events in the OAB interval can be recognized in this section. The existing proposal of Ch. timorensis as a proxy for the boundary is also valid, as the taxonomy, morphometric variation, and stratigraphic range of this species are now much better understood than when it was first proposed as a proxy. The global range of this species is also beneficial, as it can be recognized in many different places (see Fig. 3 in Grădinaru, 2022) and would allow the boundary to be correlated widely. Ch. timorensis first occurs in the Deşli Caira section just before the beginning of the peak of a positive carbon isotope excursion but below the first occurrences of Anisian ammonoids and a concurrent palaeomagnetic reversal. Therefore, if Ch. timorensis were to be chosen as the proxy for the OAB, several ammonoids traditionally considered to be Olenekian would now range into the Anisian and ammonoid zones may end up straddling the boundary. Further work is planned on the remaining unpublished conodont samples from the section, on potential new sampling of parts of the section where the stratigraphic position of archival samples is uncertain, and on a more detailed study to delimit morphotypes of Ch. timorensis at Deşli Caira and elsewhere.
Research data are stored on Geological Survey of Canada databases and can be accessed by contacting the primary author.
The author has declared that there are no competing interests.
Publisher's note: Copernicus Publications remains neutral with regard to jurisdictional claims made in the text, published maps, institutional affiliations, or any other geographical representation in this paper. While Copernicus Publications makes every effort to include appropriate place names, the final responsibility lies with the authors.
Thanks must first go to Valentin Paraschiv, who graciously gave up his time to guide the author to the Deşli Caira section, and to Iuliana Lazar and Antoneta Seghedi for their help with logistics and communication. Dana Caudle assisted with sample collection in the field. The samples were processed by Hillary Taylor and picked by Jane Broatch. Discussions on conodont taxonomy and the geology and palaeontology of the Deşli Caira section with Eugen Grădinaru and Mike Orchard proved to be invaluable for the production of this paper. Reviews by Marco Balini, two anonymous reviewers, and the editors Luke Mander and Francesca Sangiorgi helped to improve the final version.
Funding for this research was provided by a Small Research Grant of The Micropalaeontological Society, whose support is gratefully acknowledged.
This paper was edited by Luke Mander and reviewed by Marco Balini and two anonymous referees.
Agematsu, S., Orchard, M. J., and Sashida, K.: Reconstruction of an apparatus of Neostrachanognathus tahoensis from Oritate, Japan and species of Neostrachanognathus from Oman, Palaeontology, 51, 1201–1211, 2008.
Arthaber, G.: Die cephalopodenfauna der reiflinger Kalke, Beitrage zur Palaontologie und Geologie Osterreich-Ungarns und des Orients, 10, 1–112, 1896.
Arthaber, G.: Die alpine Trias des Mediterran-Gebietes, Lethaea Geognostica, II, Das Mesozoicum I, 224–391, 1906.
Assereto, R.: Die Binodosus-Zone. Ein Jahrhundert wissenschaftlicher Gegensätze, Sitzungsberichte der Österreichische Akademie der Wissenschaften, 179, 25–53, 1971.
Assereto, R.: Aegean and Bithynian: Proposal for two new Anisian substages, in: The Stratigraphy of the Alpine-Mediterranean Triassic, Schriftenreihe der erdwissenschaftlichen Komission, edited by: Zapfe, H., Österreichische Akadamie der Wissenschaften, 2, 23–39, 1974.
Assereto, R., Jacobshagen, V., Kauffmann, G. and Nicora, A.: The Scythian/Anisian boundary in Chios, Greece, Rivista Italiana di Paleontologia e Stratigrafia, 85, 715–735, 1980.
Atudorei, V.: Constraints on the Upper Permian to Upper Triassic marine carbon isotope curve. Case studies from the Tethys, Unpublished PhD Thesis, University of Lausanne, Switzerland, 160 pp., 1999.
Atudorei, V., Baud, A., Crasquin, S., Galbrun, B., Grădinaru, E., Mirăuţă, E., Maurice, R., and Zerrari, S.: Extended scientific report of the Peri-Tethys project, in: The Triassic of North Dobrogea, Lausanne, edited by: Baud, A., Geological Museum, 64 pp., 1997.
Balini, M., Lucas, S. G., Jenks, J. F., and Spielmann, J. A.: Triassic ammonoid biostratigraphy: an overview, in: The Triassic Timescale, edited by: Lucas, S. G., Geol. Soc. Lond. Spec. Publ., 334, 221–262, 2010.
Balini, M., Lăcătuş, A., Grădinaru, E., and Lazăr, I.: Towards the definition of the GSSP of the Anisian Stage (Middle Triassic): the ammonoid fauna of Deşli Caira section and its correlation with Chios (Greece) and Kçira (Albania), in: The 14th Romanian Symposium on Palaeontology, 14–15 September 2023, Bucharest, 21 pp., 2023.
Balini, M., Lăcătuş, A., Grădinaru, E., and Lazăr, I.: Aegean ammonoids from the Anisian (Middle Triassic) GSSP Candidate Section Deşli Caira (Northern Dobrogea, Romania), Rivista Italiana di Paleontologia e Stratigrafia, 130, 711–759, 2024.
Baltres, A.: Pre-Cenomanian, Mesozoic lithostratigraphic units from North Dobrogea, Studii şi Cercetări de Geologie (2003, published in 2005), 48, 49–90, 2005.
Baltres, A., Mirăuţă, E., Panin, N., and Ghenea, C.: Geological Map Romania, Agighiol Sheet 135d, scale 1:50 000, Geological Institute of Romania, 2020.
Bender, H.: Zur Gliederung der Mediterranen Trias II. Die Conodontenchronologie der Mediterranen Trias, Annales Géologique Pays Hellénique, 19, 465–540, 1970.
Brühwiler, T., Goudemand, N., Galfetti, T., Bucher, H., Baud, A., Ware, D., Hermann, E., Hochuli, P. A., and Martini, R.: The Lower Triassic sedimentary and carbon isotope records from Tulong (South Tibet) and their significance for Tethyan palaeoceanography, Sediment. Geol., 222, 314–332, 2009.
Bucher, H.: Lower Anisian Ammonoids from the northern Humboldt Range (northwestern Nevada, USA) and their bearing upon the Lower-Middle Triassic boundary, Eclog. Geolog. Helv., 82, 945–1002, 1989.
Chen, A.-F., Zhang, Y., Golding, M. L., Wu, H.-T., and Liu, J.-Q.: Upper Changhsingian to lower Anisian conodont biostratigraphy of the Datuguan section, Nanpanjiang Basin, South China, Palaeogeogr. Palaeocl. Palaeoecol., 616, 111470, https://doi.org/10.1016/j.palaeo.2023.111470, 2023.
Chen, Y., Jiang, H., Ogg, J. G., Zhang, Y., Gong, Y., and Yan, C.: Early-Middle Triassic boundary interval: Integrated chemo-bio-magneto-stratigraphy of potential GSSPs for the base of the Anisian Stage in South China, Earth Planet. Sc. Lett., 530, 115863, https://doi.org/10.1016/j.epsl.2019.115863, 2020.
Chen, Y. L., Twitchett, R. J., Jiang, H. S., Richoz, S., Lai, X. L., Yan, C. B., Sun, Y. D., Liu, X. D., and Wang, L. N.: Size variation of conodonts during the Smithian–Spathian (Early Triassic) global warming event, Geology, 41, 823–826, 2013.
Chen, Y. L., Jiang, H. S., Lai, X. L., Yan, C. B., Richoz, S., Liu, X. D., and Wang, L. N.: Early Triassic conodonts of Jiarong, Nanpanjiang Basin, southern Guizhou Province. South China, J. Asian Earth Sci., 105, 104–121, 2015.
Chen, Y. L., Richoz, S., Krystyn, L., and Zhang, Z. F.: Quantitative stratigraphic correlation of Tethyan conodonts across the Smithian-Spathian (Early Triassic) extinction event, Earth-Sci. Rev., 195, 37–51, 2019.
Chen, Z. Q. and Benton, M. J.: The timing and pattern of biotic recovery following the end-Permian mass extinction, Nat. Geosci., 5, 375–383, 2012.
Chhabra, N. L. and Sahni, A.: Late Lower Triassic and early Middle Triassic conodont faunas from Kashmir and Kumaun sequences in Himalaya, J. Paleontol. Soc. India, 25, 135–147, 1981.
Crasquin-Soleau, S. and Grădinaru, E.: Early Anisian ostracode fauna from the Tulcea Unit (Cimmerian North Dobrogean Orogen, Romania), Annales de Paléontologie, 82, 59–116, 1996.
Dagys, A. A.: Lower Triassic Conodonta from the Southeastern Pamirs: fossil plates and explanatory text, in: Trias Sibiri, edited by: Dagys, A. S., Trudy Instituta Geologii i Geofiziki (Novosibirsk), 767, 73–89, 1990.
Duan, J. Y.: Permian–Triassic conodonts from southern Jiangsu and adjacent areas, with indexes of their colour alteration, Acta Micropalaeontol. Sin., 4, 351–368, 1987.
Echevarría, J. and Ros-Franch, S.: Biogeographic response to major extinction events: The case of Triassic bivalves, Palaeogeogr. Palaeoclim. Palaeoecol., 638, 112053, https://doi.org/10.1016/j.palaeo.2024.112053, 2024.
Fantini Sestini, N.: Lower Anisian (Aegean) Ammonites from Chios Island (Greece), Rivista Italiana di Paleontologia e Stratigrafia, 87, 41–66, 1981.
Forel, M. B. and Grădinaru, E.: A unique diversity hotspot for Polycopidae (Ostracoda) in the Triassic of North Dobrogea, Hist. Biol., 34, 995–1018, 2021.
Friesenbichler, E., Hautmann, M., Grădinaru, E., and Bucher, H.: A highly diverse bivalve fauna from a Bithynian (Anisian, Middle Triassic) Tubiphytes-microbial buildup in North Dobrogea (Romania), Pap. Palaeontol., 7, 447–495, 2021a.
Friesenbichler, E., Hautmann, M., and Bucher, H.: The main stage of recovery after the end-Permian mass extinction: Taxonomic rediversification and ecologic reorganization of marine level-bottom communities during the Middle Triassic, Peer J., 9, e11654, https://doi.org/10.7717/peerj.11654, 2021b.
Gaetani, M.: Anisian/Ladinian boundary field workshop, Southern Alps-Balaton Highlands, 27 June–4 July 1993, Albertiana, 12, 5–9, 1993.
Gaetani, M., Jacobshagen, V., Nicora, A., Kauffmann, G., Tselepidis, V., Fantini Sestini, N., Mertmann, D., and Skourtsis-Coroneou, Y.: The Early-Middle Triassic boundary at Chios (Greece), Rivista Italiana di Paleontologia e Stratigrafia, 98, 181–204, 1992.
Galfetti, T., Bucher, H., Brayard, A., Hochuli, P. A., Weissert, H., Kuang, G. D., Atudorei, V., and Guex, J.: Late Early Triassic climate change: insights from carbonate carbon isotopes, sedimentary evolution and ammonoid paleobiogeography, Palaeogeogr. Palaeoclim. Palaeoecol., 243, 394–411, 2007.
Galfetti, T., Bucher, H., Martini, R., Hochuli, P. A., Weissert, H., Crasquin-Soleau, S., Brayard, A., Goudemand, N., Brühwiler, T., and Kuang, G. D.: Evolution of Early Triassic outer platform paleoenvironments in the Nanpanjiang Basin (South China) and their significance for the biotic recovery, Sediment. Geol., 204, 36–60, 2008.
Garzanti, E., Jadoul, E., Nicora, A., and Berra, E.: Triassic of Spiti (Tethys Himalaya, N India), Rivista Italian di Paleontologia e Stratigrafia, 101, 267–300, 1995.
Germani, D.: New data on ammonoids and biostratigraphy of the classical Spathian Kçira sections (Lower Triassic, Albania), Rivista Italiana di Paleontologia e Stratigrafia, 103, 267–292, 1997.
Golding, M. L.: Early Anisian (Middle Triassic) conodonts from Romania and China, with comments on their role in the recognition and correlation of the base of the Anisian, J. Earth Sci., 32, 573–591, 2021.
Golding, M. L. and Kılıç, A. M.: Reconstruction of the multielement apparatus of the conodont Gladigondolella tethydis (Huckriede) using multivariate statistical analysis; implications for taxonomy, stratigraphy, and evolution, Rivista Italiana di Paleontologia e Stratigrafia, 130, 1–18, 2024.
Goudemand, N., Orchard, M. J., Bucher, H., and Jenks, J.: The elusive origin of Chiosella timorensis (Conodont Triassic), Geobios, 45, 199–207, 2012.
Grădinaru, E.: Mesozoic rocks in North Dobrogea: an overview, Field Guidebook, Central and North Dobrogea, Romania, 1–4 October 1995, IGCP Project No. 369, Comparative Evolution of PeriTethyan Rift Basins, 17–28, 1995.
Grădinaru, E.: Workshop on the Lower-Middle Triassic (Olenekian-Anisian), 7–10 June 2000, Tulcea, Romania, in: Conference and Field Trip. Field Trip Guide, Bucharest, 37 pp., 2000.
Grădinaru, E.: Triassic-Jurassic boundary events in North Dobrogea (Romania) as recorded in basinal marine environments, in: 5th Field Workshop of IGCP 458, Transdanubian Range (Hungary) and Northern Calcareous Alps (Austria), Poster, 5–10 September 2005, 2005.
Grădinaru, E.: The Olenekian–Anisian/Early–Midddle Triassic boundary, and assessment of the potential of conodonts for chronostratigraphic calibration of the Triassic timescale, Acta Palaeontologica Romaniae, 18, 3–51, 2022.
Grădinaru, E. and Gaetani, M.: Upper Spathian to Bithynian (Lower to Middle Triassic) brachiopods from North Dobrogea (Romania), Rivista Italiana di Paleontologia e Stratigrafia, 125, 91–123, 2019.
Grădinaru, E. and Ivanova, D.: Foraminiferal biostratigraphy across the Olenekian-Anisian boundary in the Deşli Caira section (Northern Dobrogea, Romania): preliminary results, in: STS/IGCP 467 Field Meeting, 5–8 September 2002, Veszprém, Hungary, 10–11, 2002.
Grădinaru, E. and Sobolev, E. S.: Ammonoid and nautiloid biostratigraphy around the Olenekian-Anisian boundary in the Tethyan Triassic of North Dobrogea (Romania): correlation with the Boreal Triassic, in: Boreal Triassic 2006, 16–19 August 2006, Longyearbyen, Svalbard, edited by: Nakrem, H. A. and Mørk, A., NGF Abstracts and Proceedings of the Geological Society of Norway, 56–58, 2006.
Grădinaru, E., Kozur, H., Nicora, A., and Orchard, M. J.: The Chiosella timorensis lineage and correlation of the ammonoids and conodonts around the base of the Anisian in the GSSP candidate at Deşli Caira (North Dobrogea, Romania), Albertiana, 34, 34–39, 2006.
Grădinaru, E., Orchard, M. J., Nicora, A., Gallet, Y., Besse, J., Krystyn, L., Sobolev, E. S., Atudorei, N.-V., and Ivanova, D.: The Global Boundary Stratotype Section and Point (GSSP) for the base of the Anisian Stage: Deşli Caira, North Dobrogea, Romania, Albertiana, 36, 54–67, 2007.
Henderson, C. M., Golding, M. L., and Orchard, M. J.: Conodont sequence biostratigraphy of the Lower Triassic Montney Formation, Bull. Can. Petrol. Geol., 66, 7–22, 2018.
Hofmann, R., Goudemand, N., Wasmer, M., Bucher, H., and Hautmann, M.: New trace fossil evidence for an early recovery signal in the aftermath of the end-Permian mass extinction, Palaeogeogr. Palaeoclim. Palaeoecol., 310, 216–226, 2011.
Horacek, M. and Grădinaru, E.: The Spathian–Anisian (Lower–Middle Triassic) Boundary in the candidate GSSP section at Deşli Caira, Romania: Review of existing data, new findings, and comparison with other candidates, Palaeogeogr. Palaeoclim. Palaeoecol., 613, 111407, 2023.
Hounslow, M. W. and Muttoni, G.: The geomagnetic polarity timescale for the Triassic: linkage to stage boundary definitions, in: The Triassic Timescale, edited by: Lucas, S. G., Geol. Soc. Lond. Spec. Publ., 334, 61–102, 2010.
Hounslow, M. W., Szurlies, M., Muttoni, G., and Nawrocki, J.: The magnetostratigraphy of the Olenekian-Anisian boundary and a proposal to define the base of the Anisian using a magnetozone datum, Albertiana, 36, 62–67, 2007.
Ishida, K. and Hirsch, F.: The Triassic conodonts of the NW Malayan Kodiang Limestone revisited: Taxonomy and paleogeographic significance, Gondwana Res., 19, 22–36, 2011.
Jacobshagen, V., Gaetani, M., Nicora, A., Tselepedis, V., Kauffman, G., Mertmann, D., Skourtsis-Coroneou, V., and Fantini Sestini, N.: The Early/Middle Triassic boundary on Chios Island: preliminary results of a reinvestigation, Bull. Geol. Soc. Greece, 28, 25–38, 1993.
Kılıç, A. M., Plasencia, P., Ishida, K., Hirsch, F., and Yümün, Z. U.: Reflections on the apparatus of Triassic Gondolellacea (Conodonta) and the question of sexually dimorphs, in: Conodonts from the Andes, Proceedings of the 3rd International Conodont Symposium, edited by: Albanesi, G. L. and Ortega, G., Asociación Paleontológica Argentina, 13, 63–67, 2013.
Kittl, E.: Beiträge zur Kenntnis der Triasbildungen der Nordöstlichen Dobrudscha. Denkschriften der Akademie der Wissenschaften, Mathematisch-Naturwissenschaftliche Klasse, 81, 447–532, 1908.
Konstantinov, A. G. and Klets, T. V.: Stage boundaries of the Triassic in Northeast Asia, Stratigr. Geol. Correlat., 17, 173–191, 2009.
Lehrmann, D. J., Stepchinski, L., Altiner, D., Orchard, M. J., Montgomery, P., Enos, P., Ellwood, B. B., Bowring, S. A., Ramezani, J., Wang, H., Wei, J., Yu, M., Griffiths, J. D., Minzoni, M., Schaal, E. K., Li, X., Meyer, K. M., and Payne, J. L.: An integrated biostratigraphy (conodonts and foraminifers) and chronostratigraphy (paleomagnetic reversals, magnetic susceptibility, elemental chemistry, carbon isotopes and geochronology) for the Permian-Upper Triassic strata of Guandao section, Nanpanjiang Basin, South China, J. Asian Earth Sci., 108, 117–135, 2015.
Leu, M., Bucher, H., Baud, A., Vennemann, T., Luz, Z., Hautmann, M., and Goudemand, N.: An expanded Smithian–Spathian (Lower Triassic) boundary from a reefal build-up record in Oman: implications for conodont taxonomy, high-resolution biochronology and the carbon isotope record, Pap. Palaeontol., 9, e1481, https://doi.org/10.1002/spp2.1481, 2023.
Liang, L., Tong, J., Song, H., Song, T., Tian, L., Song, H., and Qiu, H.: Lower-Middle Triassic conodont biostratigraphy of the Mingtang section, Nanpanjiang Basin, South China, Palaeogeogr., Palaeoclim. Palaeoecol., 459, 381–393, 2016.
Maekawa, T., Komatsu, T., Shigeta, Y., Takashima, R., and Yamaguchi, T.: Carbonate carbon isotope chemostratigraphy and conodont biostratigraphy around the Smithian–Spathian boundary in the Panthalassan carbonate succession (SW Japan), J. Asian Earth Sci., 205, 1–11, https://doi.org/10.1016/j.jseaes.2020.104570, 2021.
Matsuda, T.: Early Triassic conodonts from Kashmir, India. Part 3: Neospathodus 2, J. Geosci. Osaka City Univ., 26, 87–110, 1983.
Matsuda, T.: Late Permian to Early Triassic condont paleobiogeography in the “Tethys Realm”, in: The Tethys: Her Paleogeography and Paleobiogeography from Paleozoic to Mesozoic, edited by: Nakazawa, K. and Dickins, J. M., Tokai University Press, Tokyo, Japan, 157–170, 1985.
Meço, S.: Conodont biostratigraphy of Triassic pelagic strata, Albania, Rivista Italiana di Paleontologia e Stratigrafia, 105, 251–266, 1999.
Meço, S.: Litho-biostratigraphy and the conodonts of Palaeozoic/Triassic deposits in Albania, Palaeontographica Abteilung A, 290, 131–197, 2010.
Mertmann, D. and Jacobshagen, V.: Upper Olenekian (Spathian) ammonoids from Chios (Lower Triassic, Greece): taxonomy and stratigraphic position, Rivista Italiana di Paleontologia e Stratigrafia, 109, 417–447, 2003.
Mirăuţă, E.: Über die Conodontenfaunen des oberen Werfens und des tieferen Anis der nördlichen Dobrudscha/Rumänien, Geologica et Palaeontologica, 8, 149–158, 1974.
Mirăuţă, E.: Conodont biostratigraphy in the Deşli Caira section, in: Workshop on the Lower-Middle Triassic (Olenekian-Anisian), 7–10 June 2000, Tulcea, Romania, Conference and Field Trip, Conference Section, Bucharest, 81–84, 2000.
Mirăuţă, E., Gheorghian, D., and Bădiceanu, M.: Données biostratigraphiques sur la Formation de Cataloi (Dobrogea de Nord, Roumanie), Romanian J. Stratigr., 75, 21–27, 1993.
Muttoni, G. and Rettori, R.: New biostratigraphic data on the Triassic of Marathovouno hillock area (Chios Island, Greece), Rivista Italian di Paleontologia e Stratigrafia, 99, 461–472, 1994.
Muttoni, G., Kent, D. V., and Gaetani, M.: The base of the Anisian. A candidate Global Stratotype Section and Point from Chios Island, Greece, Albertiana, 13, 37–43, 1994.
Muttoni, G., Kent, D. V., and Gaetani, M.: Magnetostratigraphy of a Lower-Middle Triassic boundary section from Chios (Greece), Phys. Earth Planet. Inter., 92, 245–260, 1995.
Muttoni, G., Kent, D. V., Meço, S., Nicora, A., Gaetani, M., Balini, M., Germani, D., and Rettori, R.: Magnetobiostratigraphy of the Spathian to Anisian (Lower to Middle Triassic) Kçira section, Albania, Geophys. J. Int., 127, 503–514, 1996.
Muttoni, G., Nicora, A., Balini, M., Katz, M., Schaller, M., Kent, D. V., Maron, M., Meço, S., Rettori, R., Doda, V., and Nazaj, S.: A candidate GSSP for the base of the Anisian from Kçira, Albania, Albertiana, 45, 39–49, 2019.
Nakazawa, K. and Bando, Y.: Lower and Middle Triassic Ammonites from Portuguese Timor (Palaeontological Study of Portuguese Timor, 4), in: Memoirs of the Faculty of Science, Series of geology and mineralogy, Kyoto University, 34, 83–114, 1968.
Nicora, A.: Lower Anisian platform – conodonts from the Tethys and Nevada: taxonomic and stratigraphic revision, Palaeontographica, 157, 88–107, 1977.
Nogami, Y.: Trias-Conodonten von Timor, Malaysien und Japan, in: Memoirs of the Faculty of Science, Series Geology and Mineralogy, Kyoto University, 34, 115–136, 1968.
Ogg, J. G.: Publication Announcement: Early-Middle Triassic boundary interval: Integrated chemo-bio-magneto-stratigraphy of potential GSSPs for the base of the Anisian Stage in South China. By Yan Chen, Haishui Jiang, James G. Ogg, Yang Zhang, Yifan Gong & Chunbo Yan, 2019. Earth and Planetary Science Letters, https://doi.org/10.1016/j.epsl.2019.115863, Albertiana, 45, 7–73, 2019.
Ogg, J. G., Huang, C., and Hinnov, L.: Triassic timescale status: a brief overview, Albertiana, 41, 3–30, 2014.
Orchard, M. J.: Taxonomy and correlation of Lower Triassic (Spathian) segminate conodonts from Oman and revision of some species of Neospathodus, J. Paleontol., 69, 110–122, 1995.
Orchard, M. J.: Multielement conodont apparatuses of Triassic Gondolelloidea, Spec. Pap. Palaeontol., 73, 73–101, 2005.
Orchard, M. J.: Triassic conodonts and their role in stage boundary definition, in: The Triassic Timescale, edited by: Lucas, S. G., Geol. Soc. Lond. Spec. Publ., 334, 139–161, 2010.
Orchard, M. J.: North American Spathian (Upper Olenekian, Lower Triassic) Neogondolellin conodonts, Pap. Palaeontol., 8, e1409, https://doi.org/10.1002/spp2.1409, 2022.
Orchard, M. J. and Golding, M. L.: The Neogondolella constricta (Mosher and Clark, 1965) group in the Middle Triassic of North America: speciation and distribution, J. Paleontol., 97, 1161–1191, https://doi.org/10.1017/jpa.2023.52, 2024.
Orchard, M. J. and Tozer, E. T.: Triassic conodont biochronology, its calibration with the ammonoid standard, and a biostratigraphic summary of the Western Canada Sedimentary Basin, in: Triassic of the Western Canada Sedimentary Basin, edited by: Moslow, T. F. and Wittenberg, J., Bull. Can. Petrol. Geol., 45, 675–692, 1997.
Orchard, M. J., Grădinaru, E., and Nicora, A.: A summary of the conodont succession around the Olenekian-Anisian boundary at Deşli Caira, North Dobrogea, Romania, in: The Global Triassic, edited by: Lucas, S. G. and Spielmann, J. A., New Mexico Museum of Natural History and Science Bulletin, 41, 341–346, 2007a.
Orchard, M. J., Lehrmann, D. J., Jiayong, W., Hongmei, W., and Taylor, H.: Conodonts from the Olenekian-Anisian boundary beds, Guandao, Guizhou Province, China, in: The Global Triassic, edited by: Lucas, S. G. and Spielmann, J. A., New Mexico Museum of Natural History and Science Bulletin, 41, 347–354, 2007b.
Ovtcharova, M., Bucher, H., Schaltegger, U., Galfetti, T., Brayard, A., and Guex, J.: New Early to Middle Triassic U–Pb ages from South China: calibration with ammonoid biochronozones and implications for the timing of the Triassic biotic recovery, Earth Planet. Sc. Lett., 243, 463–475, 2006.
Ovtcharova, M., Goudemand, N., Hammer, Ø., Kuang, G. D., Cordey, F., Galfetti, T., Schaltegger, U., and Bucher, H.: Developing a strategy for accurate definition of a geological boundary through radio-isotopic and biochronological dating: the Early-Middle Triassic boundary (South China), Earth-Sci. Rev., 146, 65–76, 2015.
Pia, J.: Grundbegriffe der Stratigraphie mit ausfiihrlicher anwendung auf die Europaische Mitteltrias, Deuticke, Leipzig, Wien, 252 pp., 1930.
Pietsch, C., Ritterbush, K. A., Thompson, J. R., Petsios, E., and Bottjer, D. J.: Evolutionary models in the Early Triassic marine realm, Palaeogeogr. Palaeoclim. Palaeoecol., 513, 65–85, 2019.
Rosenberg, G.: Geleitworte zu den Tabellen der Nord- und Südalpinen Trias der Ostalpen, Jahrbuch der Geologischen Bundesanstalt, 102, 477–479, 1959.
Sebe, O.-G., Crasquin, S., and Grădinaru, E.: Early and Middle Anisian deep-water ostracods (Crustacea) from North Dobrogea (Romania), Revue de Paléobiologie, 32, 509–529, 2013.
Simionescu, J.: Studii geologice si paleontologice din Dobrogea. III. Fauna tirasică dela Deşli-Caïra, Academia Română Publicaţiunile Fondului Vasile Adamachi, 465–493, 1910.
Simionescu, J.: Studii geologice si paleontologice din Dobrogea. V. Fauna tirasică inferioară din Dobrogea, Academia Română Publicaţiunile Fondului Vasile Adamachi, 63–77, 1911.
Song, H. J., Wignall, P. B., Tong, J. N., Song, H. Y., Chen, J., Chu, D. L., Tian, L., Luo, M., Zong, K. Q., Chen, Y. L., Lai, X. L., Zhang, K. X., and Wang, H. M.: Integrated Sr isotope variations and global environmental changes through the Late Permian to early Late Triassic, Earth Planet. Sc. Lett., 424, 140–147, 2015.
Stone, J.: Review of investigative techniques used in the study of conodonts, in: Conodonts: Investigative Techniques and Applications, British Micropalaeontological Society Series, edited by: Austin, R. L., Ellis Horwood Ltd., 17–34, 1987.
Summesberger, H. and Wagner, L.: Der Stratotypus des Anis (Trias), Annalen des Naturhistorischen Museums in Wien, 1972, 515–538, 1972.
Sun, Y. D., Joachimski, M. M., Wignall, P. B., Yan, C. B., Chen, Y. L., Jiang, H. S., Wang, L. N., and Lai, X. L.: Lethally hot temperatures during the Early Triassic greenhouse, Science, 338, 366–370, 2012.
Sweet, W. C.: Uppermost Permian and Lower Triassic conodonts of the Salt Range and Trans-Indus Ranges, West Pakistan, in: Stratigraphic Boundary Problems, Permian and Triassic of West Pakistan, edited by: Kummel, B. and Teichert, C., Special Publication 4, Department of Geology, University of Kansas, 207–275, 1970.
Tong, J., Chu, D., Liang, L., Shu, W., Song, H., Song, T., Song, H., and Wu, Y.: Triassic integrative stratigraphy and timescale of China, Sci. China Earth Sci., 62, 189–222, 2019.
Waagen, W. and Diener, C.: Untere Trias, in: Entwurf einer Gliederung der pelagischen Sedimente des Trias-Systems, edited by: von Mojsisovics, E., Waagen, W., and Diener, C., Sitzungsberichte Akademie Wissenschaft Wien, Mathematische – naturwissenschaftliche Klasse, 104, 1271–1302, 1895.
Wang, H., Wang, X., Li, R., and Wei, J.: Triassic conodont succession and stage subdivision of the Guandao section, Bianyang Luodian, Acta Palaentol. Sin., 44, 611–626, 2005.
Wignall, P. B.: The Worst of Times: How Life on Earth Survived Eighty Million Years of Extinctions, Princeton University Press, 2015.
Wu, S., Golding, M. L., Zhao, H., Han, C., Zhang, X., Chen, Z.-Q., and Lyu, Z.: Lower to Middle Triassic conodont biostratigraphy and carbonate carbon isotope chemostratigraphy of the Zuodeng area, Guangxi, South China, and its relevance for stratigraphic correlation from the Induan to the Anisian, Palaeogeogr. Palaeoclim. Palaeoecol., 636, 111965, https://doi.org/10.1016/j.palaeo.2023.111965, 2024.
Wu, S., Golding, M. L., Han, C., Lyu, Z., and Zhao, L.: Morphometric analysis of the conodont Chiosella timorensis (Nogami) from the early Anisian of Romania and China, and its significance for the definition of the Olenekian-Anisian Boundary, Palaeontology, in press, 2025.
Zakharov, Y. D. and Rybalka, S. V.: A standard for the Permian-Triassic in the Tethys, in: Problems of the Permian and Triassic Biostratigraphy of East USSR, edited by: Zakharov, Y. D. and Unoprienko, Y. I., USSR Academy of Science, Far Eastern Scientific Centre, Vladivostock, 6–48, 1987.
Zapfe, H. (Ed.): Trias in Österreich, in: Die Stratigraphie der alpin-mediterranen Trias. Schriftenreihe der Erdwissenschaftlichen Kommissio-nen, Österreichische Akademie der Wissenschaften, 2, 245–251, 1974.
Zapfe, H. (Ed.): Das Forschungsproject “Triassic of the Tethys Realm” (IGCP Proj. 4). Abschlußbericht, in: Neue Beiträge zur Biostratigraphie der Tethys-Trias, Schriftenreihe der Erdwissenschaft-lichen Kommissionen, Österreichische Akademie der Wissenschaften, 5, 7–16, 1983.