the Creative Commons Attribution 4.0 License.
the Creative Commons Attribution 4.0 License.
 | 17 Jan 2018
| 17 Jan 2018
A humid early Holocene in Yemen interpreted from palaeoecology and taxonomy of freshwater ostracods
Munef Mohammed
Peter Frenzel
Dietmar Keyser
Fadhl Hussain
Abdulkareem Abood
Abdulmajed Sha'af
Sadham Alzara'e
Sakher Alammari
Lake or marsh sediments in the Qa'a Jahran–Dhamār area indicate a period of higher moisture availability in the early Holocene of the highlands of Yemen. Forty-two marl–peat sediment samples from eight stratigraphic sections of that area have been collected and are examined for the first time for their ostracod associations. Eight species belonging to seven genera and four families are reported. Their ecological tolerances and preferences are used to investigate the climatic and environmental changes in the early to mid-Holocene. Our data are compared and correlated with previous archaeological results, particularly from the region of Qa'a Jahran (Dhamār) in the vicinity of the village of Beyt Nahmi. We conclude that the wettest period of the Holocene was from about 7900 to 7400 cal yr BP, when northwards incursion of the Indian Ocean Monsoon caused intensified monsoon precipitation over southern Arabia.
- Article
(6002 KB) - Full-text XML
-
Supplement
(452 KB) - BibTeX
- EndNote





Multidisciplinary research including geology, geomorphology, ecology, archaeology and anthropology in different parts of the world has addressed Holocene climatic changes and the development of linked geomorphologic features such as lakes (e.g. Pachur and Hoelzmann, 1991; de Menocal et al., 2000; Mischke, 2001; Fleitmann and Matter, 2009; Broxton et al., 2011; Mischke et al., 2012). Some of these studies reported the distribution and development of Holocene palaeolakes in the Arabian Peninsula, where water availability is a challenge for the future (e.g. Acres, 1982; Wilkinson, 1997; Lézine et al., 1998; Fleitmann et al., 2003; Davies, 2006; Parker et al., 2006; Rosenberg et al., 2011; Engel et al., 2012; Enzel et al., 2015). Understanding changes in the moisture availability of the past is a prerequisite for a sustainable management of water resources. The few previous surveys which have been done in Yemen, to review the relationship between the early human settlements and Holocene climatic changes, recognized lacustrine deposits and extensive scatters of shells of freshwater molluscs in the plain of Qa'a Jahran at very high elevations (2400 m above sea level) and some deserted districts in Ramlat as-Sab'atayn and Wadi Juba in Marib (Acres, 1982; Wilkinson, 1997; Lézine et al., 1998; Davies, 2006). These studies illustrate that the Yemeni inland desert and highlands experienced strong climatic changes leading to the development and disappearance of freshwater lakes. The presence of dried palaeolakes in the Dhamār highlands is one of the striking geomorphological features in Yemen; however, little is known of their history and distribution. The deposits that are exposed in the quarries and shallow dry wells dug by local people can be considered excellent evidence for the formation of such lakes in tectonic depressions. The fluctuations of both moisture and aridity during the Holocene are the major factors affecting the development of lakes in the Arabian Peninsula (Mayewski et al., 2004).
Ostracoda are small crustaceans characterized by having a bivalve shell hinged along the dorsal margin. Most species are of microscopic size. Their shell is composed of low-magnesium calcite, may be smooth or ornamented, and fossilizes well. The rich ostracod fossil record is not only due to the fossils' ease of preservation but also due to their high adaptability to different environmental conditions. Ostracods are sensitive to fluctuations of ecological parameters mainly at the water–sediment interface; these parameters can be temperature, salinity, pH, oxygen, turbulence or trophic level amongst other factors. The record of these variations can be observed at several levels: abundance, diversity, species composition and morphological variability (Carbonel et al., 1988). Furthermore, shell chemistry, i.e. stable isotope and trace element signatures, provides additional information about past environmental conditions.
Only a few papers have been published on freshwater ostracods of Yemen. Malz (1976) discussed the changes in carapace morphology and the taxonomic problems of the Holocene genera Heterocypris Sars and Cyprinotus Brady and their fossil relative Cheikella Sohn and Morris, 1963 described from Cenozoic freshwater deposits in Saudi Arabia. His study also includes some specimens from Yemen. The work by Dumont et al. (1986) focused on the taxonomy and distribution of Cladocera, Copepoda and Ostracoda from freshwaters of South Yemen. Three valves of Cyprinotus rostrata Lowndes have been discovered by Munef Mohammed (unpublished data) from Anthropocene dry mud of the city of Aden. Mazzini and Sardella (2004) found some freshwater ostracods during their study of the Quaternary of Socotra Island. Mohammed et al. (2013) studied the taxonomy and distribution of Holocene freshwater ostracods from the northern part of Socotra Island. Taking into consideration this poor knowledge of Quaternary Ostracoda from Yemen, our study contributes to the record of Ostracoda biodiversity.
The present study aims to investigate the taxonomy and palaeoecology of freshwater ostracods extracted from marl–peat sediments of the Holocene palaeolake deposits of the Qa'a Jahran–Dhamār highlands. It will provide a contribution to the knowledge of climate change during the Holocene.
Yemen is situated at the south-western corner of the Arabian Peninsula framed by the Gulf of Aden and the Red Sea. Four main physiographic provinces can be recognized: the coastal plain province, the high volcanic mountains province, the Hadhramaut–Mahara Plateau province and the desert terrain province (Rub Al-Khali). The study area is an inland intermountain plain within the high volcanic mountains province (Al-Rawi, 2008). This province is characterized by the occurrence of Cenozoic (Yemen trap series) and Quaternary (Yemen volcanic series) volcanoes. The main volcanic activity in the study area is related to the Dhamār–Rada volcanic field, an extensively faulted and fractured graben and half graben area, probably related to the Red Sea rifting. The late Cenozoic and Quaternary volcanic activities (about 5 Ma ago) were confined to the area around Dhamār (Geukens, 1966; Mattash et al., 2013) and are represented in the study area by the volcanic mountains of Isbil and Al-Lisi. Archaeological and historical studies support the record of continuous Quaternary volcanic activity almost to the present day. These studies provide information on the volcanic history of the region and its effects on human populations, e.g. through the exploitation of obsidian sources in the past (Khalidi et al., 2010).
The NW–SE-oriented Qa'a Jahran plain (Fig. 1) is a semi flat area surrounded by high plateaus rising approximately 500+ m (Davies, 2006). It is situated in a graben trough related to the tectonic regime of the opening Red Sea. Quaternary alluvium and lake deposits varying in thickness were deposited on the top of this plain (Davies, 2006). Runoff channels from upslope and intermittent valleys which run along the tectonic zones are mostly dry stream beds and only flood during the high rainy seasons. The region is a semi-arid steppe with annual rainfall less than 300 mm. It receives limited moisture twice yearly in the form of winter north-westerly circulation and summer monsoonal rains (Parker et al., 2006; Davies, 2006).
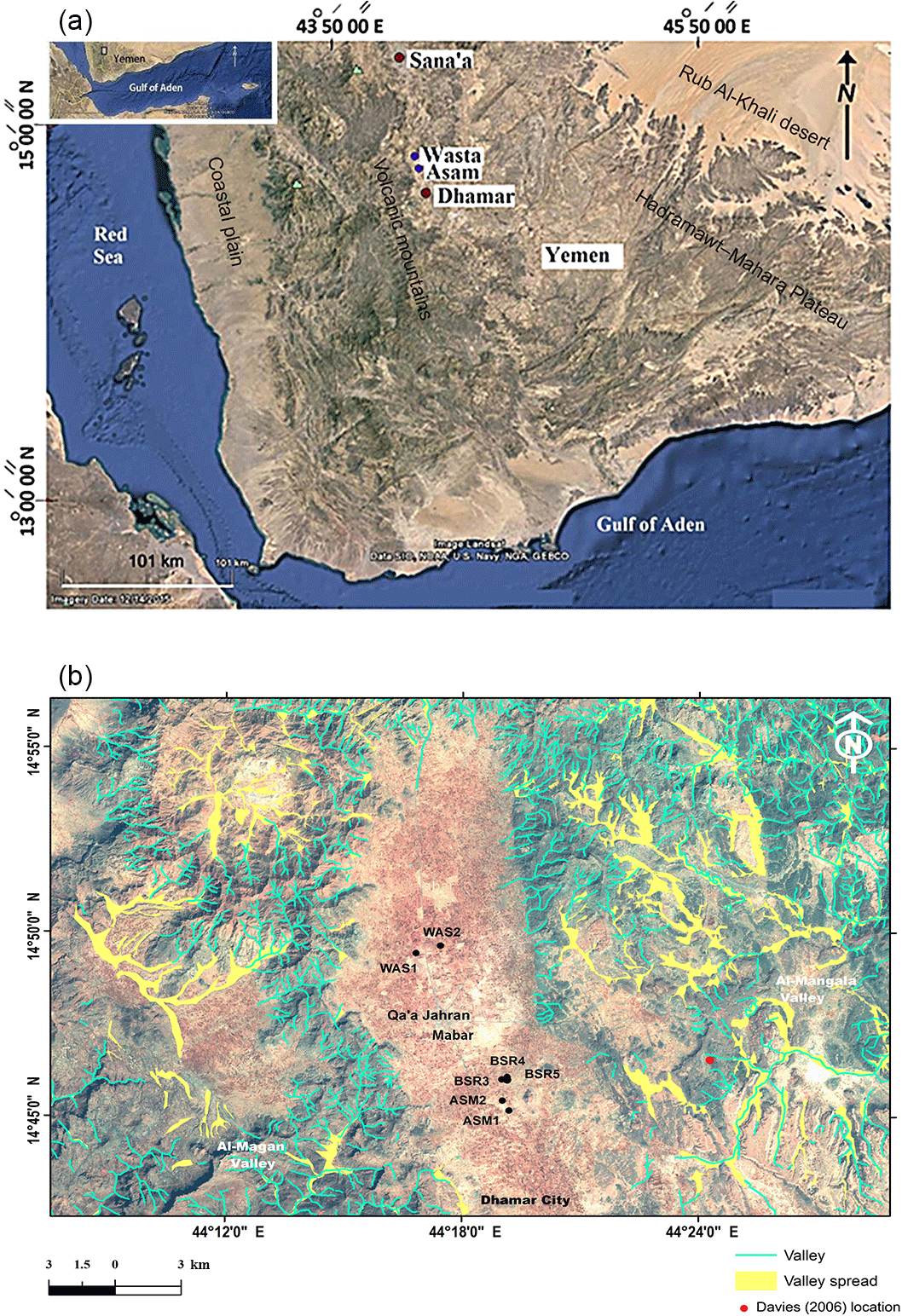
Figure 1Study area in Yemen (a) and studied sections on a Landsat ATM + image of Qa'a Jahran (b).

Wilkinson (1997) and Davies (2006), in their investigations of the Holocene lacustrine deposits of the Dhamār highlands, reported that these semi-arid valleys responded to past changes in atmospheric circulation by the rapid development of lakes and marshes during wet times and the expansion of soils during drier phases. They deduced that the alternating development of highland lakes and soils resulted from the shifts in Holocene palaeoclimate due to fluctuations in the Indian Ocean Monsoon between moist and dry phases.
Two field trips were made to the Qa'a Jahran–Dhamār area during November 2013 and March 2014 to collect the material used in the current study. Grey to dark grey marl layers were traced laterally through eight exposed sections in the northern part of the basin of Qa'a Jahran. The layers under investigation show similarity in lithology but differ in thickness. They are overlain by a modern agricultural horizon of fine sandy silt loam and underlain by light grey marl deposits.
Forty-three marl samples of variable thickness were collected from those eight sections from eight shallow wells dug by villagers (Figs. S1 and S2 in the Supplement). They were mostly taken at intervals of 10–20 cm focusing on the darker layers. The sampling locations along with geographical coordinates and the deposit descriptions are given in Table 1. All the samples were subject to standard micropalaeontological techniques (Moore, 1961). A unit weight of 250 g (ca. 5 cm thick) of fresh unweathered sediment sample was always taken. Samples were covered in a pan with 29 % hydrogen peroxide to separate the mud and clay from the shells. After allowing a sample to soak for about 12 h, water was added and boiled. The samples were washed with running tap water over a 200 mesh sieve (opening of 0.074 mm) to remove mud-size sediment. About 10 to 20 g of washed residue of sandy sediment, shells and rock fragments were extracted from each sample. The ostracods were collected by using a 00 brush under a stereomicroscope, placed into micro-slides and identified. No ostracods with complete soft parts were found. Representatives of the recorded ostracods were sent to Hamburg, Jena and Rome. Scanning electron microscope (SEM) photos were taken at the Zoological Institute and Museum at Hamburg University, the Institute of Zoology at Jena University and the Istituto di Geologia Ambientale e Geoingegneria Roma. Specimens of all ostracod species were deposited in the Department of Earth and Environmental Sciences, Faculty of Science, Sanaa University. All measurements are given in millimetres. Statistical analysis using the program package PAST (Hammer et al., 2001) was done to support the interpretation of the palaeoenvironments. The ostracods collected in this study are in the collections of the first author except for the depicted material that is deposited in Hamburg, Jena and Rome.
The systematics are based on Moore (1961), Hartmann and Puri (1974), Meisch (2000), and Karanovic (2012). References within the synonymy lists are selected based on taxonomic, biogeographic and ecological significance. The abbreviations used are as follows: RV – right valve; LV – left valve; C – carapace; V – valve; L – length; H – height; ASM – Asam sections; WAS – Wasta sections; BSR – Beyt Rashed sections.
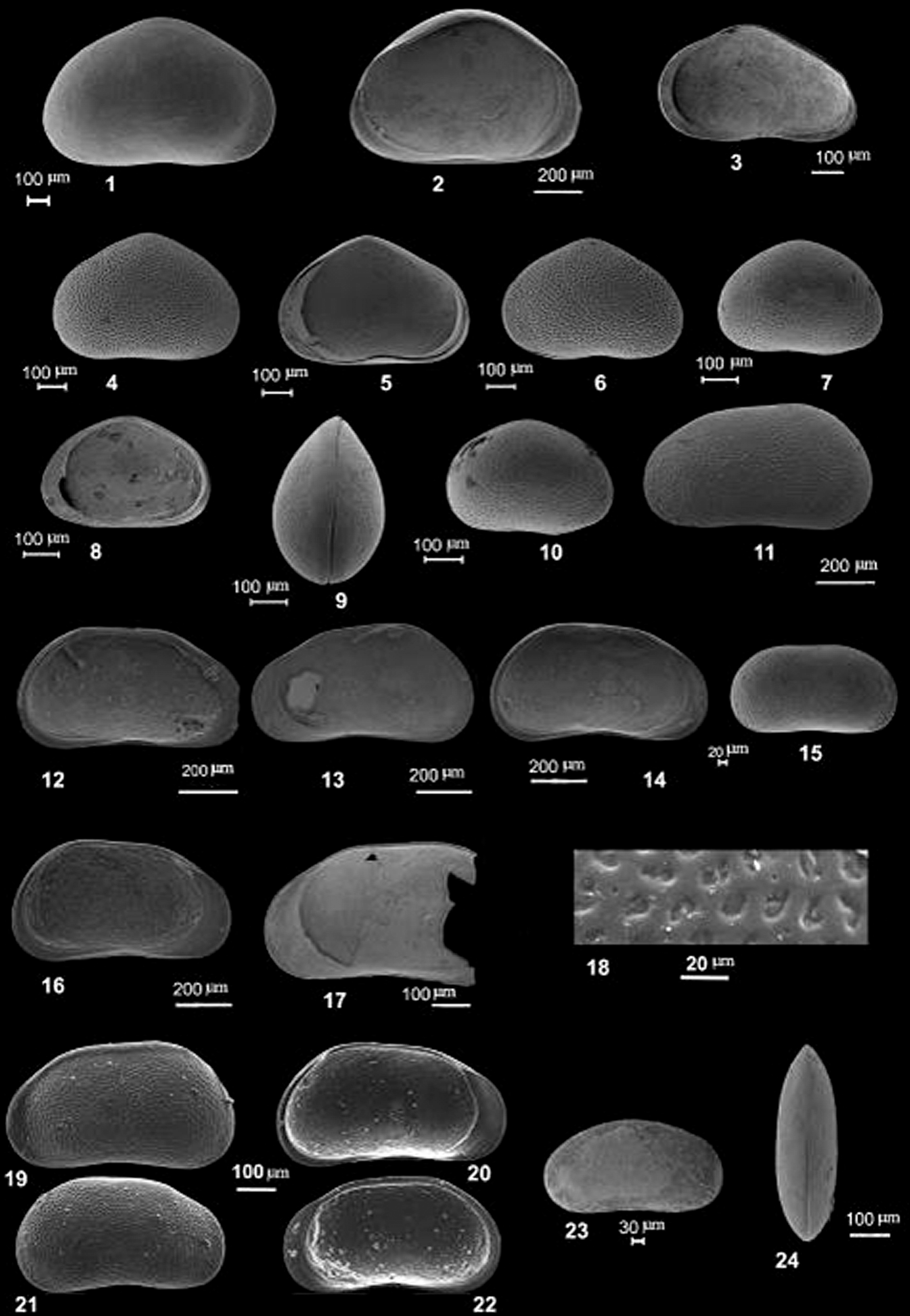
Plate 1(1–3) Heterocypris salina (Brady, 1868): (1) RV, external lateral view, section ASM1, sample 1; (2) LV, internal lateral view, section WAS2a, sample 2; (3) Juv., RV, internal lateral view, section ASM1, sample 1. (4–6) Sarscypridopsis aculeata (Costa, 1847): (4) LV, external, lateral view, section ASM1, sample 1; (5) RV, internal, lateral view, section ASM1, sample 1; (6) LV, external, lateral view, section ASM1, sample 2. (7–10) Cypridopsis concolor Daday, 1900: (7) RV, external, lateral view, section ASM1, sample 2; (8) RV, internal, lateral view, section ASM1, sample 1; (9) Carapace, dorsal view, section ASM1, sample 1; (10) RV, external, lateral view, section ASM1, sample 1. (11–22) Pseudocandona cf. albicans (Brady, 1864): (11) LV, external, lateral view, section WAS2, sample 2; (12) LV, internal, lateral view, section WAS2, sample 2; (13) RV, internal, lateral view, section WAS2a, sample 2; (14) LV, internal, lateral view, section WAS2, sample 2; (15) RV, juv., external, lateral view, section ASM1, sample 1; (16) LV, internal, lateral view, section WAS2, sample 2; (17) RV, internal, lateral view, section WAS2, sample 2; (18) Detail of surface ornamentation, RV, section WAS2, sample 2; (19) LV, external, lateral view, WAS2, sample 2; (20) LV, internal, lateral view, section WAS2, sample 1; (21) Juv., RV, external, lateral view, section WAS2, sample 2; (22) Juv., RV, internal, lateral view, section WAS2, sample 2. (23–24) Fabaeformiscandona cf. breuili (Paris, 1920): (23) Carapace, RV, external lateral view, section WAS2a, sample 2; (24) Carapace, dorsal view, section WAS2a, sample 3.
Family Cyprididae Baird, 1845
Subfamily Cyprinotinae Bronstein, 1947
Genus Heterocypris Claus, 1892
Heterocypris salina (Brady, 1868)
(Plate 1, figs. 1–3)
*1868 Cypris salina Brady: 368, pl. 28, figs. 8–13.
1980 Hemicypris posterotruncata Bate; McClure & Swain: pl. 2, fig. 6.
1996 Heterocypris salina (Brady); Schöning: 41–42, figs. 1–4, 7–9.
2000 Heterocypris salina (Brady); Meisch: 354, fig. 148A–G.
2001 Heterocypris salina (Brady); Griffiths et al.: 763.
2004 Heterocypris salina (Brady); Rosenfeld et al.: 173, pl. 1, fig. 15.
2008 Heterocypris salina (Brady); Beker et al.: 18, 19, pl. 3, figs. 6–8.
2010 Heterocypris cf. salina (Brady); Mischke & Almogi-Labin: fig. 9.
2011 Heterocypris salina (Brady); Rosenberg et al.: Supplementary data 33–34 [partim non Heterocypris salina].
2012 Heterocypris salina (Brady); Mischke et al.: pl. 2, figs. 7–10, 18.
2012 Heterocypris salina (Brady); Fuhrmann: 228, pl. 108, figs. 1a–d, 2a–d.
2014 Heterocypris salina (Brady); Kalbe et al.: fig. 3j.
2015 Heterocypris salina (Brady); Mischke et al.: 157.
2016 Heterocypris salina (Brady); Kalbe et al.: fig. 6g–h.
Material: Adults: 8 RV, 9 LV; juveniles: 25 RV, 20
LV.
Size: RV: L –
1.13 mm; H – 0.70 mm. LV: L – 1.2 mm; H –
0.77 mm.
Occurrence:
ASM1, WAS2a.
Geographical distribution: Holarctic with
introductions into the Southern Hemisphere (Meisch, 2000);
Anthropocene sediments from Sudan
(Schöning, 1996); Holocene from Jordan (Mischke et al., 2012) and Israel
(Flako-Zaritsky et al., 2011; Mischke et al.,
2014).
Remarks: A
large number of the specimens recorded in the present study are juvenile
carapaces and valves.
Subfamily Cypridopsinae Kaufmann, 1900
Tribe Cypridopsini Bronstein, 1947
Genus Sarscypridopsis McKenzie, 1977
Sarscypridopsis aculeata (Costa, 1847)
(Plate 1, figs. 4–6)
*1847 Cypris aculeata Costa: 11–12, pl. 3, fig. 5.
2000 Sarscypridopsis aculeata (Costa); Meisch: 392, fig. 163A–D.
2001 Sarscypridopsis aculeata (Costa); Griffiths et al.: 763.
2012 Sarscypridopsis aculeata (Costa); Karanovic: 422, fig. 4d, fig. 135e–f, fig. 137d.
2012 Sarscypridopsis aculeata (Costa); Fuhrmann: 248, pl. 118, fig. 1a–d.
Material: Adults: 13 C, 23 RV, 22 LV; juveniles: 16 RV, 12 LV.
Size: RV: L – 0.70 mm; H – 0.43 mm. LV: L – 0.66–0.69 mm; H – 0.44–0.45 mm.
Occurrence: ASM1, WAS2a.
Geographical distribution: A cosmopolitan species (Meisch, 2000).
Genus Cypridopsis Brady 1867
Cypridopsis concolor Daday, 1900
(Plate 1, figs. 7–10)
*1900 Cypridopsis vidua var. concolor Daday: 190, pl. 30a–c.
2000 Cypridopsis vidua concolor Daday; Meisch: 372.
2012 Cypridopsis concolor Daday; Fuhrmann: 238, pl. 113, fig. 2a–d.
Material: 4 C, 4 RV, 2 LV.
Size: RV: L – 0.48 mm; H – 0.31 mm. LV: L – 0.45 mm; H – 0.30 mm,
W – 0.32 mm.
Occurrence: ASM1, WAS2a.
Geographical distribution: Budapest in Hungary; Negorci wetland in
Macedonia; fossil from Pleistocene deposits in central Germany.
Remarks: The present study follows Fuhrmann (2012) in separating C. concolor from C. vidua because
of the differences in size, the outline of the carapace and the design of
the postero-internal border zone of the left valve.
Family Candonidae Kaufmann, 1900
Subfamily Candoninae Kaufmann, 1900
Tribe Candonini Kaufmann, 1900.
Group Compressa
Genus Pseudocandona Kaufmann, 1900
Pseudocandona cf. albicans (Brady, 1864)
(Plate 1, figs. 11–22)
*1864 Candona albicans Brady:. 61, pl. 4, figs. 6–10.
1968 Candona albicans Brady; Bhatia: 471, pl. 2, fig. 4a–c.
1973 Pseudocandona albicans (Brady); Danielopol: 235.
1999 Pseudocandona albicans (Brady); Mazzini et al.: pl. 2, fig. 6.
?2001 Pseudocandona albicans (Brady); Griffiths et al.: 762.
2011 Pseudocandona albicans (Brady); Özulug: 95, fig. 2.
?2012 Pseudocandona parallela (Müller 1900); Fuhrmann: 86, pl. 37, figs. 1a–e, 2a–e.
Material: Adults: 18 RV, 17 LV; juveniles: 6 C, 11 RV, 12 LV.
Size: LV: L – 0.7 mm; H – 0.5 mm.
Occurrence: ASM1, WAS2, WAS2a.
Geographical distribution: Probably Holarctic (Meisch, 2000).
Remarks: There is some confusion about the identification of the current
species because its morphology closely resembles a number of taxa belonging
to the Candonini group. Our adult specimens are ornamented with dense pits
corresponding to the original description of Candona albicans by Brady (1864); however,
later literature reported smooth carapaces from different regions of the
world and regarded Brady's specimens as juveniles. The pitted adult valves
of P. albicans documented in the present study may indicate a response of the animals
to some specific environmental factor(s).
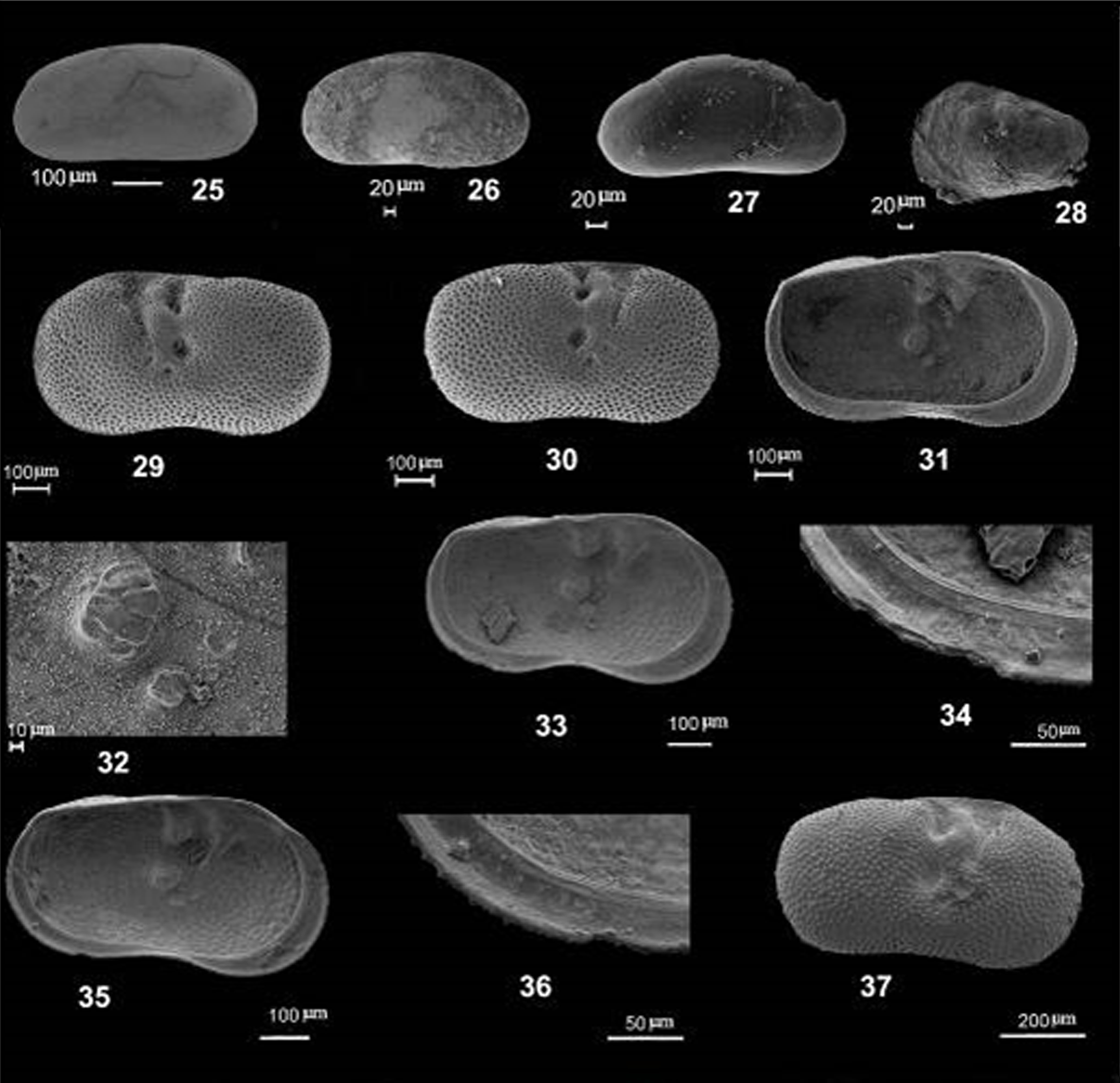
Plate 2(25–26) Fabaeformiscandona cf. breuili (Paris, 1920): (25) Carapace, LV, external lateral view, section ASM1, sample 1; (26) Carapace, LV, external lateral view, section ASM1, sample 1. (27) ?Fabaeformiscandona sp., juv., RV, external lateral view, ASM1, sample 2. (28) Leucocythere sp., juv., LV, external, lateral view, section ASM1, sample 2. (29–37) Ilyocypris bradyi (Ramdohr, 1808): (29) LV, external, lateral view, section ASM1, sample 1; (30) RV, external, lateral view, section ASM1, sample 1; (31) LV, internal, lateral view, section ASM1, sample 1; (32) Muscle scars, LV. ASM1, sample 1; (33) LV, internal, lateral view, section WAS2a, sample 2; (34) Postero-ventral marginal area of LV, internal, lateral view, WAS2a, sample 2; (35) LV, internal, lateral view, WAS2a, sample 2; (36) Postero-ventral marginal area of LV, internal, lateral view, WAS2a, sample 2; (37) RV, external, lateral view, WAS2a, sample 2.
Genus Fabaeformiscandona Krstic, 1972
Fabaeformiscandona cf. breuili (Paris, 1920)
(Plate 1, figs. 23, 24; Plate 2, figs. 25, 26)
1920 Candona breuili Paris: 477, pl. 18 figs. 1–3.
2000 Fabaeformiscandona breuili (Paris); Meisch: 135, figs. 56A–C.
2012 Fabaeformiscandona breuili (Paris); Fuhrmann: 46, pl. 17, figs. 1a–f, 2a–b, 3a–b.
Material: 5 C.
Size: LV: L – 0.46 mm; H – 0.23 mm.
Occurrence: ASM1, WAS2, WAS2a.
Geographical distribution: Germany, France and the Czech Republic.Remarks: The present species resembles Fabaeformiscandona breuili (Paris, 1920) in lateral outline and
the overlapping of valves, but it is smaller in size.
?Fabaeformiscandona sp.
(Plate 2, fig. 27)
Material: 1 juvenile LV.
Size: LV: L – 0.26 mm; H – 0.12 mm.
Occurrence: ASM1.
Remarks: The single left valve of a juvenile of this species was not enough
to identify it precisely.
Family Ilyocyprididae Kaufmann, 1900
Genus Ilyocypris Brady & Norman, 1889
Ilyocypris bradyi Sars, 1890
(Plate 2, figs. 29–37)
*1890 Ilyocypris bradyi Sars: 59.
2000 Ilyocypris bradyi Sars; Meisch: 253, figs. 107A–C.
2001 Ilyocypris bradyi Sars; Griffiths et al.: 762.
2012 Ilyocypris bradyi Sars; Mischke et al.: pl. 2, fig. 23–25.
2012 Ilyocypris bradyi Sars; Fuhrmann: 150, pl. 69, figs. 1a–f, 2a–d.
2014 Ilyocypris bradyi Sars; Kalbe et al.: fig. 3e–f.
2015 Ilyocypris cf. bradyi Sars; Mischke et al.: figs 7–15.
2015 Ilyocypris bradyi Sars; Kalbe et al.: fig. 7m–n.
Material: Adults: 30 RV, 27 LV, juveniles: 21 RV, 23 LV.
Size: RV: L – 0.80 mm; H – 0.42 mm. LV: L – 0.84 mm; H – 0.45 mm.
Occurrence: ASM1, WAS2, WAS2a, BSR4.
Geographical distribution: Holarctic (Meisch, 2000).
Remarks: There are several problems in the identification of I. bradyi because of
the intraspecific variability of its characters and the very close morphological
features with other representatives of the genus. Based on descriptions of
modern animals with soft parts and analysing the morphology of the valves,
van Harten (1979), Janz (1994) and Mazzini et al. (2014) provided important clues
about the morphological structures of the carapaces of some abundant Holocene
species and discussed the assumption that the marginal ripplet structure on
the inner lamella of Ilyocypris left valves is of primary taxonomic value. The
position of the marginal ripplets on the inner lamella in conjunction with
the inner or outer list is used here to differentiate between I. bradyi and the
non-tuberculate I. gibba.
Family Limnocytheridae Klie, 1938
Genus Leucocythere Kaufmann, 1892
Leucocythere sp.
(Plate 2, fig. 28)
Material: 1 juvenile LV.
Size: LV: L – 0.27 mm; H – 0.18 mm.
Occurrence: ASM1.
Remarks: This single juvenile left valve was not sufficient to be identified
to the species level.
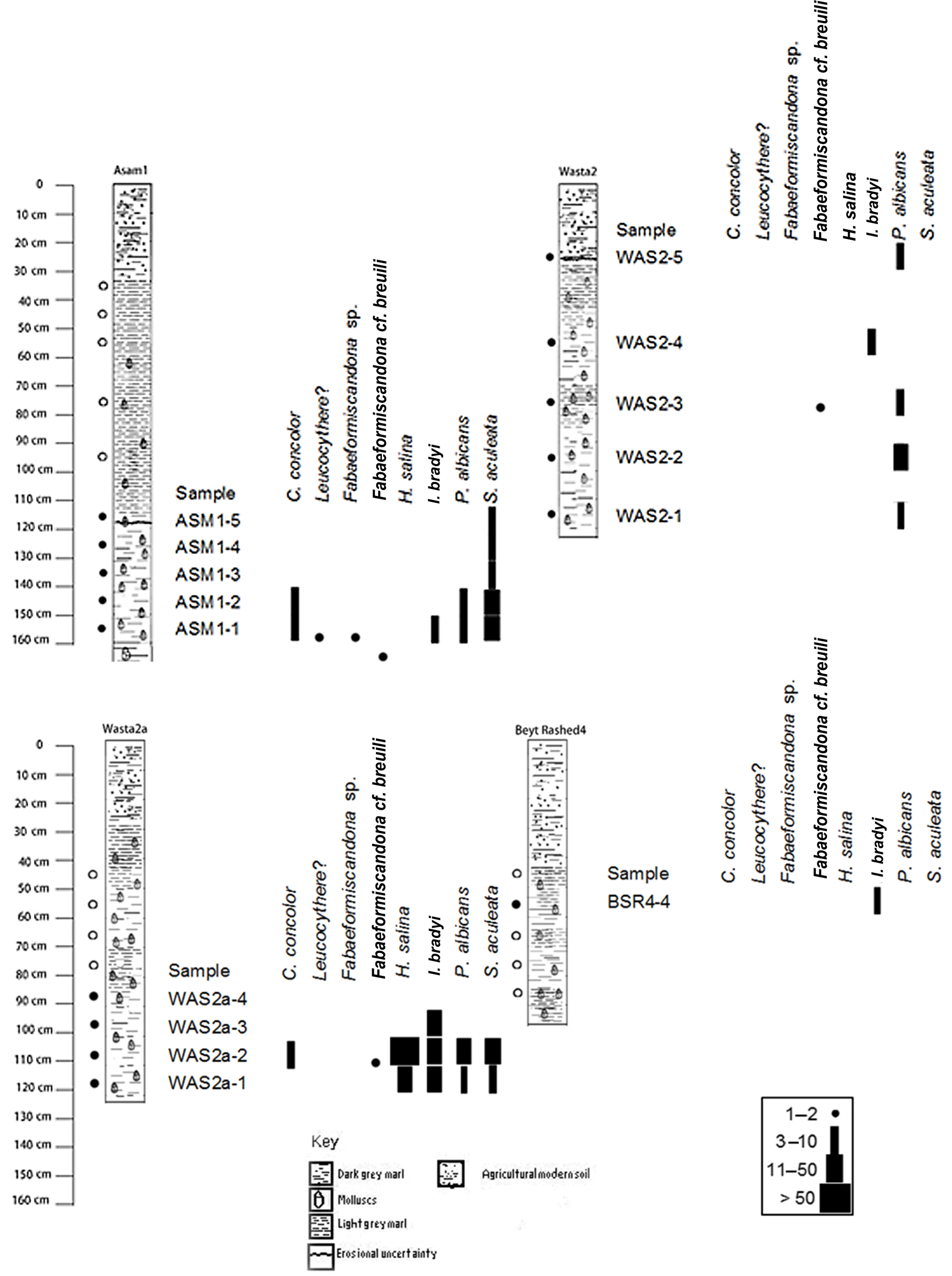
Ostracod shells were found in 15 samples belonging to four sections (ASM1, WAS2, WAS2a and BSR4) associated with molluscs and small numbers of charophyte oospores. However, a large amount of ostracod shell debris was recorded in almost all the studied samples. Levels with increased coarse grains and/or mollusc shell fragments were considered as representing an erosional unconformity. The different thicknesses of the studied sections is related to the varying morphology of the basin. The distribution of ostracods along the studied sections is illustrated in Fig. 2.
Many ecological observations on living Ostracoda can be readily applied to fossil ostracods especially when dealing with the same or closely related taxa (Martens, 1994). Information on the ecology of the species encountered in the current study has been gathered from several previous contributions (Hiller, 1972; Hartmann and Hiller, 1977; Meisch, 2000; Frenzel et al., 2010; Fuhrmann, 2012). The freshwater taxa Leucocythere sp. and ?Fabaeformiscandona sp. are not included in the following discussion of palaeoecology because of their uncertain species attribution and scarcity.
Fabaeformiscandona breuili (Paris, 1920)
Poorly known: Reported from drainage pipes, springs, caves and the interstitial groundwater.
Heterocypris salina (Brady, 1868)
Salinity: 0.4–8.6 ‰, oligohaline to low
mesohaline. Animals may occur even in pure freshwater
habitats.Temperature:
Thermoeuryplastic, 16–22 ∘C. Ganning (1971) explained that the
species prefers habitats that are high in nutrients (eutrophic) and have low
temperature and found that no animals survived for longer than 3 days at
30 ∘C.
Water depth and energy: Frequent occurrence in pools along the coast (Ganning,
1971) and in salinized ponds or streams on the mainland (Klie, 1938),
mesorheophilic (frequently found in flowing waters with various
velocities).
Habitat and substrate: Permanent and temporary ponds; sediment and phytal,
nectobenthic.
O2: Low oxygen,
< 1 mL L−1.
Life history: In ephemeral waterbodies,
Heterocypris salina needs 40 days to reach the adult stage (Ganning,
1971).
Ilyocypris bradyi Sars, 1890
Salinity: 0–4.5 ‰, freshwater to oligohaline.
Temperature: Polythermophilic, 0.1–25 ∘C.
Water depth and energy: Very shallow, mesorheophilic; it prefers slowly
flowing, cooler waters of springs, streams and ponds fed by springs (Mischke,
2001).
Habitat and substrate: Springs, ponds, swamps and estuaries, and temporary
waters. The animals recorded in lakes are usually reported as discharged by
nearby springs; endobenthic. Often in running water.
O2: Probably high oxygen > 2.5 mL L−1.
Life history: The species produces two generations annually.
Pseudocandona cf. albicans (Brady, 1864)
Salinity: < 6.3 ‰, freshwater to low mesohaline
(optimum salinity 5.5 ‰).
Temperature: Mesothermophilic, 2–24 ∘C.
Water depth and energy: Very shallow; mesorheophilic, frequently found in
flowing waters with various velocities; however, it prefers stagnant and slow
flowing waters.
Habitat and substrate: Lagoons and estuaries, swamps, ponds and lakes,
temporary waters; muddy bottom and phytal.
O2: Low oxygen, > 0 mL L−1.
Sarscypridopsis aculeata (Costa, 1847)
Salinity: 0.5–17 ‰; oligohaline to mesohaline (optimum
salinity 17 ‰). It prefers slightly brackish small waterbodies of both inland and coastal type, where it often produces large
populations, the species is rare in freshwater.
Temperature: Thermoeuryplastic, 3–25 (26) ∘C.
Water depth and energy: Very shallow, mesorheophilic (frequently found in
flowing waters with various velocities).
Habitat and substrate: Temporary and permanent ponds, sand and phytal,
nectobenthic.
O2: Low oxygen, < 1 mL L−1.
Life history: Development of juveniles is very rapid; sexual maturity is
reached within 30 days (Mischke, 2001).
Cypridopsis concolor Daday, 1900
No details on the ecological requirements of the species have been found in the literature. It was recorded from freshwater of lowland wetlands in Europe. It prefers smaller and probably also temporary warm waters (Melovski et al., 2013; Fuhrmann, 2012).
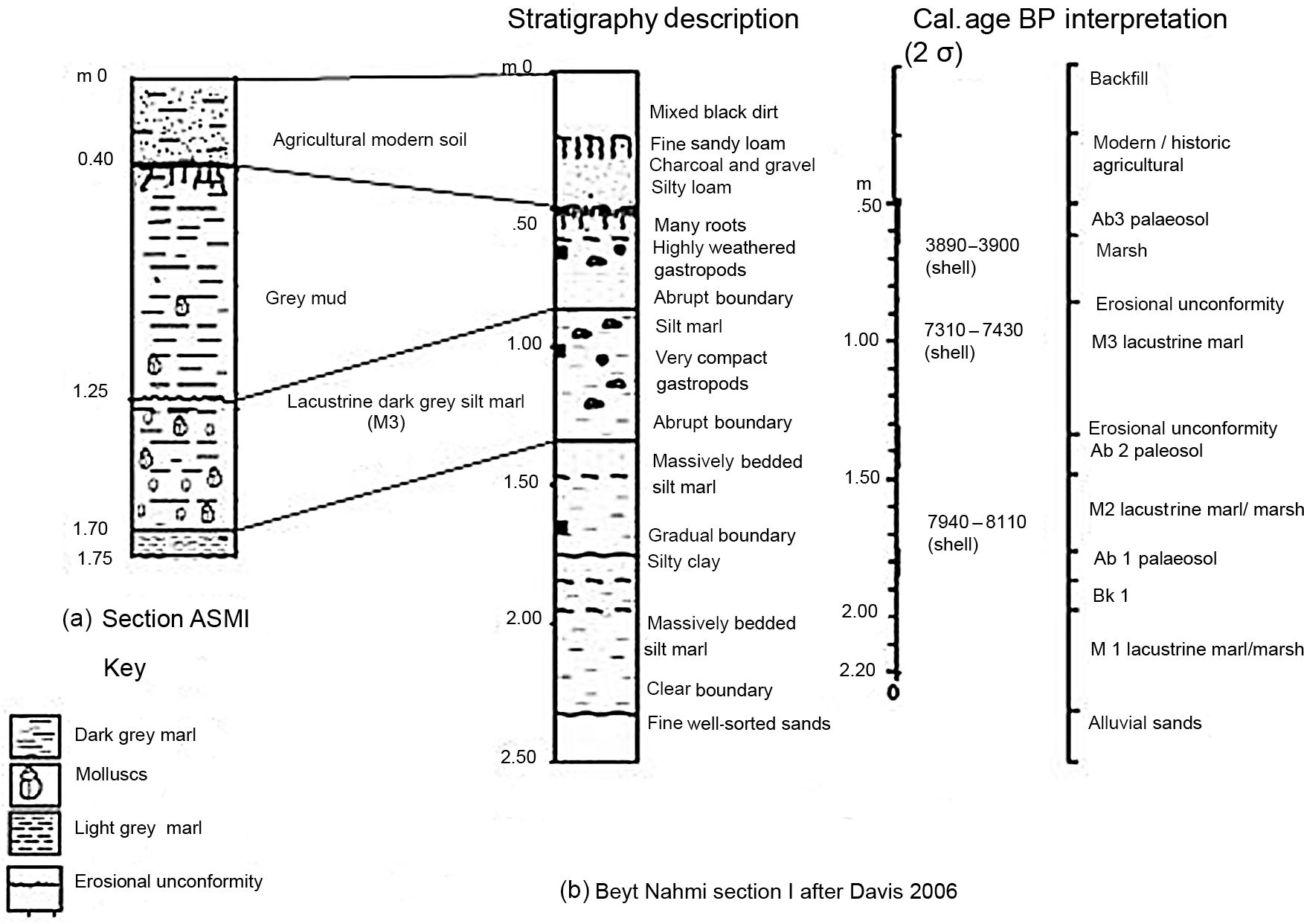
Figure 3Stratigraphic correlation between (a) present study ASM1 section and (b) Beyt Nahmi “section 1” of Davies (2006).
In her study site located outside the village of Beyt Nahmi, Davies (2006) found a sediment sequence of 2.50 m thickness from a deep trench composed of three discrete marl horizons alternating with three humus-rich horizons or palaeosols. They represent the northern margin of early to mid-Holocene deposits of lacustrine origin and marshes. Based on the location of sites, lithology and stratigraphic position, the thickest section ASM1 of the current study can be correlated to the Beyt Nahmi “section 1” of Davies (2006), which is about 4 km from the present study location (Fig. 1). This enabled us to find the levels extending from about 1.40 to 0.50 m between the grey calcareous silt marl (M3) and the uppermost palaeosol (Ab3), corresponding to levels of the grey to dark grey marl (1.70 to 0.40 m) of the ASM1 section. We could therefore use the 14C dates of “section 1” to correlate ages of about 7940 to 3900–3690 cal yr BP for the studied stratigraphic units (Fig. 3).
At the ASM1 section, ostracod associations are composed of Cypridopsis concolor, Fabaeformiscandona cf. breuili, ?Fabaeformiscandona sp., Ilyocypris bradyi, Heterocypris salina, Leucocythere sp., Pseudocandona cf. albicans and Sarscypridopsis aculeata. The most dominant species in the assemblage is S. aculeata, which has been recorded between 1.7 and 1.3 m, suggesting conditions of slightly higher salinity. I. bradyi and C. concolor are recorded with less abundance between 1.70 and 1.40 m, reflecting fresh to oligohaline conditions. The rare occurrence of P. cf. albicans instars, H. salina, Leucocythere sp. and Fabaeformiscandona sp. is attributed to post-mortem transport from nearby habitats. F. cf. breuili may indicate a groundwater flow through the lake. The darker grey sediments at the lowermost part of the section (1.7 to 1.5 m), which includes S. aculeata in higher abundance, is considered to reflect saline water (up to mesohaline) and low-oxygen conditions of a shallow lake. However, the co-occurrence of the mesorheophilic and freshwater species I. bradyi and C. concolor with the stagnant saline water-loving S. aculeata could be due to the mixing of different waterbodies.
The northern section WAS2a contains C. concolor, F. cf. breuili, H. salina, I. bradyi, P. cf. albicans and S. aculeata. It is dominated by the freshwater to oligohaline species I. bradyi. H. salina, S. aculeata and P. cf. albicans. Only a few individuals of C. concolor have been recorded in this section. The assemblage points to fresh-oligohaline conditions, probably near the shoreline or inlet of the lake since the discharge of streams frequently lowers salinity. Evidence of transportation is provided by the abundance of I. bradyi, which prefers slowly flowing water. The occurrence of H. salina and P. cf. albicans in this habitat is reasonable because they tolerate a wide range of salinity. Furthermore, evidence of short periods of increased evaporation could be indicated by the mesohaline-preferring S. aculeata, which is recorded from the lowermost samples (1.2 to 1.1 m) of the section.
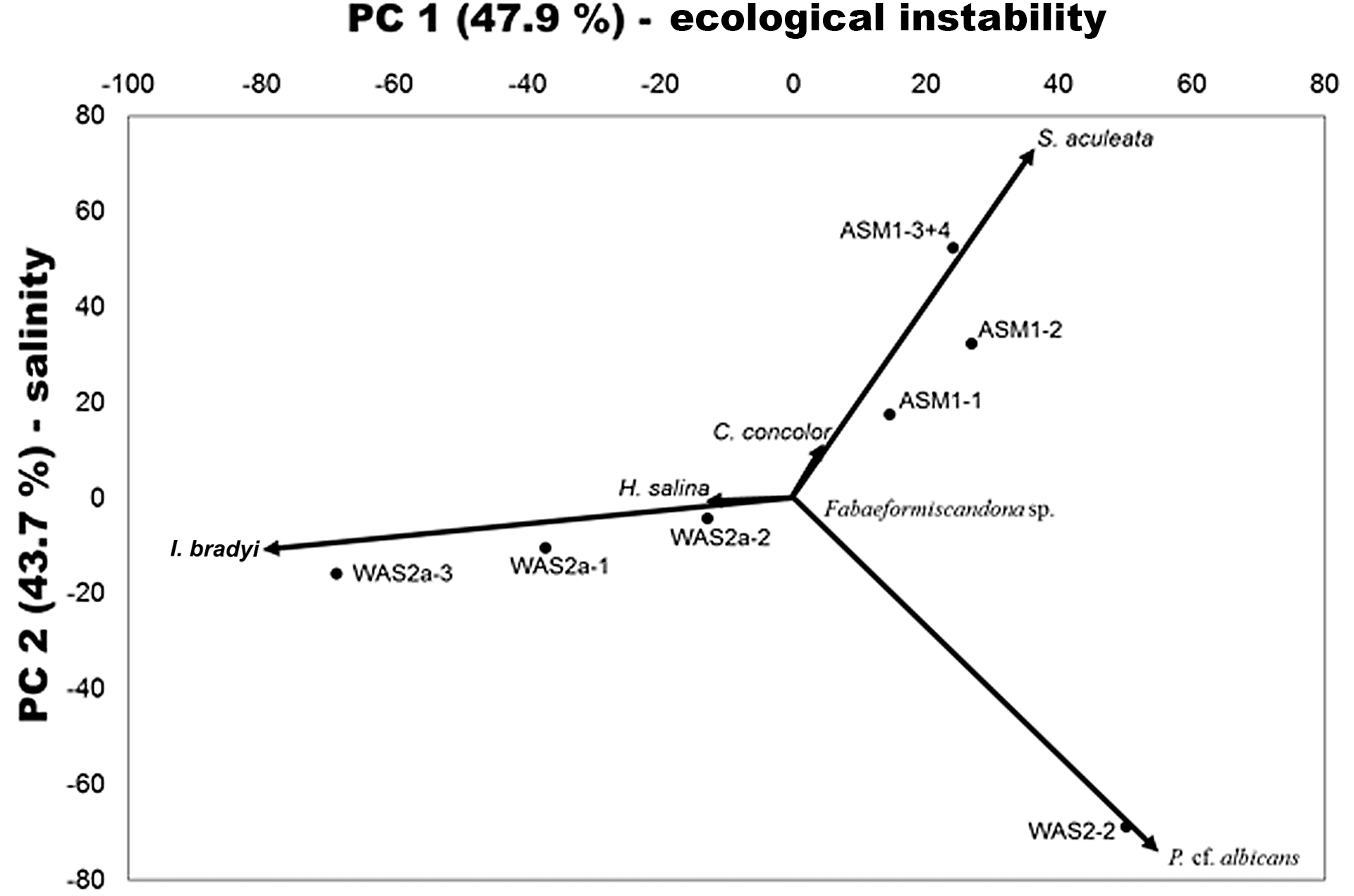
Figure 4Principal component analysis of relative ostracod abundances in samples of at least 20 counted valves from all studied sections. The percentage values of the axes show how much variance they explain. The top axis represents ecological instability, e.g. temporary vs. permanent waterbodies; the left-hand one mirrors salinity. All loadings of taxa are enlarged 100-fold for better visibility.
At section WAS2, the ostracod association displays lesser diversity than in the nearby section WAS2a. Shells of P. cf. albicans were found more often than the only other species, I. bradyi. The characters of this ostracod association indicate oligohaline waters in a small isolated habitat.
The three sections of Beyt Rashed (BSR3, BSR4, BSR5) are generally barren of Ostracoda, except at 0.6 m of BSR4, in which a few valves of I. bradyi were encountered. This may indicate the development of a small pond at the eastern shore of the lake with only a discontinuous supply of water.
The statistical analysis of ostracod distribution over all sections (Fig. 4) shows highest relative abundances of S. aculeata and P. cf. albicans and lowest negative values for I. bradyi along the top axis. Because I. badyi is a species of permanent waters and because S. aculeata as well as many Pseudocandona species often occur in temporary waters (Meisch, 2000), we assume the top axis to mirror ecological instability, i.e. temporary vs. permanent habitats. The left-hand axis reflects salinity as the high loadings of S. aculeata and its lonely presence in those samples indicate. This species tolerates salinities of up to 17, which is the highest salinity tolerance of the documented species (Frenzel et al., 2010). Considering the position of samples within the principle component analysis, a low salinity of probably fresh to oligohaline conditions is assumed for the lower and middle levels of the sections (WAS2, WAS2a and BSR4) and an elevated salinity of probably mesohaline to oligohaline conditions at ASM1. Furthermore, a trend to higher salinity and temporary waters is visible in the arrangement of samples of section ASM1 reflecting the shallowing, disintegration and finally drying out of the waterbodies in an arid climate towards the middle Holocene.
In general, the environment was a shallow fragmented lake or ponds, which could only be formed during a period of humid climate. These levels correspond to the grey silt marl (M3), which was interpreted by Davies (2006) to indicate shallow lake conditions developed during the wettest period of the Holocene ranging from 7940 to 7310–7430 cal yr BP. The uppermost part of the sections reflects a period of strong aridity. Enzel et al. (2015) regarded the sediment successions of Davies (2006) as being wetland deposits.
Several studies such as de Menocal et al. (2000), Mischke (2001), Engel et al. (2012) and Enzel et al. (2015) reported the wet period of the Holocene in a wider area including Arabia, Africa and Asia and discussed the solar cycle effects on Indian monsoon climate. Our record fits a moist period documented for the early to mid-Holocene (Fleitmann and Matter, 2009).
Eight cosmopolitan ostracod species belonging to seven genera and four families have been recorded from the lower to middle levels of Holocene stratigraphic sections of the Qa'a Jahran basin–Dhamār highlands. The palaeoenvironment was inferred relying on the ecology of their modern representatives, in order to reconstruct the history of a Holocene palaeolake. The ostracod assemblages from the Qa'a Jahran sections indicate a very shallow lake or several ponds formed during the wettest period during the early Holocene.
Northward incursion of the Indian Ocean Monsoon due to increased solar radiation caused intensified monsoon precipitation over southern Arabia during the early to mid-Holocene. The morphological setting of the studied area led to the formation of wetlands, which provided habitats for many types of plants and animals. Wetlands were distributed unevenly throughout the area because of variations in geomorphology and sources of water. The fragmented lake became more and more unstable and saline over time (from oligohaline to mesohaline) owing to the reduction in precipitation due to the retreat of the Indian Ocean Monsoon. However, the grey marl sediments which extend to the upper levels of the studied sections may indicate less wet conditions persisting for a longer period of time.
Geoarchaeological investigations on the atmospheric processes and human activity during the humid period of the Holocene in southern Arabia and in the Dhamār highlands indicate Neolithic populations.
Specimens of all ostracod species were deposited in the department of Earth and Environmental Sciences, Faculty of Science, Sana'a University, under the title “First paper of Qa'a Jahran ostracods”, abbreviation “Qaj, Qa'a Jahran”, repository of the material.
The supplement related to this article is available online at: https://doi.org/10.5194/jm-37-167-2018-supplement.
The authors declare that they have no conflict of interest.
Our thanks are due to Ilaria Mazzini (Roma) for SEM micrographs and for
helpful discussion. The work of Peter Frenzel was supported by the German
Research Foundation (DFG FR1489/5-1). We thank Antje Schwalb (TU
Braunschweig) for material from Mundafan for comparison and Anna Pint
(University of Cologne) for unpublished data from the same area. We thank
Ilaria Mazzini and Claude Meisch for their constructive
reviews.
Edited by: Alan Lord and Sev
Kender
Reviewed by: Ilaria Mazzini and one anonymous referee
Acres, B. D.: Soil classification and correlation in the Montane plains, Yemen Arab Republic: Report of Project Record, No. 72, 1982.
Al-Rawi, F.: Isostatic conditions in southern and eastern Yemen and their important geologic implications, Iraqi Journal of Science, 49, 116–123, 2008.
Baird, W.: Arrangement of the British Entomostraca, with a list of species,particularly noticing those which have as yet been discovered within the bounds of the Club, Transactions of the Berwickshire Naturalists' Club, 2, 145–158, 1845.
Beker, K., Tunoğlu, C., and Ertekin, I. K.: Pliocene-Lower Pleistocene Ostracoda Fauna from İnsuyu Limestone (Karapınar-Konya/CentralTurkey) and its Paleoenvironmental Implications, Geological Bulletin of Turkey, 51, 1–31, 2008.
Bhatia, S. B.: Pleistocene Ostracodes from the Upper Karewas of Kashmir, India, Micropaleontology, 14, 465–483, 1968.
Brady, G. S.: On species of Ostracoda new to Britain, Annales of the Magazine of Natural History, 13, 59–64, 1864.
Brady, G. S.: A synopsis of the Recent British Ostracoda, The Intellectual Observer, 12, 110–130, 1867.
Brady, G. S.: A monograph of the recent British Ostracoda, Transactions of the Linnean Society of London, 26, 353–495, 1868.
Brady, G. S. and Norman, A. M.: A monograph of the marine and freshwater Ostracoda of the North Atlantic and of northwestern Europe. I: Podocopa. Scientific Transactions of the Royal Dublin Society, 4, 63–270, 1889.
Bronstein, Z. S.: Crustacés. Ostracodes des eaux douces, Fauna SSSR Zoologicheskjy Institut Akademii Nauk SSSR, 31, 1–339, 1947.
Broxton, W. B., Mark, B. A., Donald, T. R., and Mathias, V.: Holocene tropical South American hydroclimate revealed from a decadally resolved lake sediment δ18O record, Earth Planet. Sc. Lett., 310, 192–202, https://doi.org/10.1016/j.epsl.2011.08.040, 2011.
Carbonel, P., Colin, J.-P., Danielopol, D. L., Löffler, H., and Neustrueva, I.: Paleoecology of limnic ostracodes: A review of some major topics, Palaeogeogr. Palaeocl., 62, 413–461, https://doi.org/10.1016/0031-0182(88)90066-1, 1988.
Claus, C.: Beiträge zur Kenntnis der Süsswasserostracoden. 1. Über den Körper- und Gliedmaßenbau der Cyprididen nebst Bemerkungen über einzelne innere Organe derselben, Arbeiten des zoologischen Institutes Wien, 10, 50–70, 147–216, 1892.
Costa, G.-O.: Fauna del Regno di Napoli, Animali Articulati, Crostacei, Entomostraci, Branchipodi, 7–12, https://doi.org/10.5962/bhl.title.63937, 1847.
Daday, E.: Ostracoda Hungariae. A Magyarorsza'gi Kagylo'sra'kok Maga'nrajza Kiadja a Magyar Tudoma'nyos Akade'mia, Budapest, 1900.
Danielopol, D. L.: Sur la morphologie des aesthetascs chez quelques ostracodes hypogés de la sous-famille des Candoninae (Cyprididae, Podocopida), Annales de Speléologie, 28, 233–245, 1973.
Davies, C. P.: Holocene paleoclimates of southern Arabia from lacustrine deposits of the Dhamar highlands, Yemen, Quaternary Res., 66, 454–464, 2006.
de Menocal, P., Ortiz, J., Guilderson, T., Adkins, J., Sarnthein, M., Baker, L., and Yarusinsky, M.: Abrupt onset and termination of the African Humid Period: rapid climate responses to gradual insolation forcing, Quaternary Sci. Rev., 19, 347–361, 2000.
Dumont, H., Maas, S., and Martens, K.: Cladocera, Copepoda and Ostracoda (Crustacea) from Fresh Waters in South Yemen, Fauna of Saudi Arabia, 8, 12–19, 1986.
Engel, M., Brückner, H., Pint, A., Wellbrock, K., Ginau, A., Voss, P., Grottker, M., Klasen, M., and Frenzel, P.: The early Holocene humid period in NW Saudi Arabia: sediments, microfossils and palaeo-hydrological modelling, Quaternary Int., 266, 131–141, https://doi.org/10.1016/j.quaint.2011.04.028, 2012.
Enzel, Y., Kushnir, Y., and Quade, J.: The middle Holocene climatic records from Arabia: Reassessing lacustrine environments, shift of ITCZ in Arabian Sea, and impacts of the southwest Indian and African monsoons, Global Planet. Change, 129, 69–91, https://doi.org/10.1016/j.gloplacha.2015.03.004, 2015.
Flako-Zaritsky, S., Almogi-Labin, A., Schilman, B., Rosenfeld, A., and Benjamini, C.: The environmental setting and microfauna of the oligohaline Timsah pond, Israel: The last remnant of the Kabara swamps, Mar. Micropaleontol., 80, 74–88, https://doi.org/10.1016/j.marmicro.2011.06.002, 2011.
Fleitmann, D. and Matter, A.: The speleothem record of climate variability in southern Arabia, CR Geosci., 341, 633–642, 2009.
Fleitmann, D., Burns, S. J., Mudelsee, M., Neff, U., Kramers, J., Mangini, A., and Matter, A.: Holocene monsoon recorded in a stalagmite from southern Oman, Science, 300, 1737–1739, 2003.
Frenzel, P., Keyser, D., and Viehberg, F. A.: An illustrated key and (palaeo) ecological primer for Postglacial to Recent Ostracoda (Crustacea) of the Baltic Sea, Boreas, 39, 567–575, https://doi.org/10.1111/j.1502-3885.2009.00135.x, 2010.
Fuhrmann, R.: Atlas quartärer und rezenter Ostrakoden Mitteldeutschlands, Altenburger Naturwissenschaftliche Forschungen, 15, 1–320, 2012.
Ganning, B.: On the ecology of Heterocypris salinus, H. incongruens and Cypridopsis aculeata (Crustacea: Ostracoda) from Baltic brackish-water rockpools, Mar. Biol., 8, 271–279, 1971.
Geukens, F.: Geology of the Arabian peninsula, Yemen, Geological Survey Professional paper, 506-B, B1-B23, 1966.
Griffiths, H. I., Schwalb, A., and Stevens, L. R.: Environmental change in southwestern Iran: The Holocene ostracod fauna of Lake Mirabad, Holocene, 11, 757–764, 2001.
Hammer, Ø., Harper, D. A. T., and Ryan, P. D.: PAST: Palaeontological statistics software package for education and data analysis, Palaeontol. Electron., 4, 567–575, https://doi.org/10.1006/qres.1998.1996, 2001.
Hartmann, G. and Puri, H. S.: Summary of neontological and paleontological classification of Ostracoda, Mitteilungen aus dem Hamburgischen Zoologischen Museum und Institut, 7, 7–73, 1974.
Hartmann, G. and Hiller, D.: Beitrag zur Kenntnis der Ostracodenfauna des Harzes und seines nördlichen Vorlandes (unter besonderer Berücksichtigung des Männchens von Candona candida), 125 Jahre Naturwissenschaftlicher Verein Goslar, 99–116, 1977.
Hiller, D.: Untersuchungen zur Biologie und zur Ökologie limnischer Ostracoden aus der Umgebung von Hamburg, Archiv für Hydrobiologie, Supplement, 40, 400–497, 1972.
Janz, H.: Zur Bedeutung des Schalenmerkmals “Marginalrippen” der Gattung Ilyocypris (Ostracoda, Crustacea), Stuttgarter Beiträge für Naturkunde, B (Geologie und Paläontologie), 206, 1–19, 1994.
Kalbe, J., Ames, C., Nowell, A., Cordova, C., and Pokines, J. T.: Pleistocene wetlands in the eastern desert of Jordan: Environmental reconstructions by ostracod and gastropod analysis, Abstracts of the “Middle Palaeolithic in the Desert II” conference, 11, 13 December 2014, Bordeaux, France, 2014.
Kalbe, J., Mischke, S., Dulski, P., and Sharon, G.: The Middle Palaeolithic Nahal Mahanayeem Outlet site, Israel: reconstructing the environment of Late Pleistocene wetlands in the eastern Mediterranean from ostracods, J. Archaeol. Sci., 54, 385–395, https://doi.org/10.1016/j.jas.2014.04.018, 2015.
Kalbe, J., Jagher, R., and Pümpin, C.: The spring of Nadaouiyeh Aïn Askar – Paleoecology of a Paleolithic oasis in arid central Syria, Palaeogeogr. Palaeocl., 446, 252–262, https://doi.org/10.1016/j.palaeo.2016.01.030, 2016.
Karanovic, I.: Recent Freshwater Ostracods of the World: Crustacea, Ostracoda, Podocopida, Springer, Heidelberg, https://doi.org/10.1007/978-3-642-21810-1, 2012.
Kaufmann, A.: Über die Gattung Acanthopus Vernet und eine neue Süsswassercytheride, Zool. Anz., 15, 393–395, 1892.
Kaufmann, A.: Cypriden und Darwinuliden der Schweiz, Rev. Suisse Zool., 8, 209–423, 1900.
Khalidi, L., Oppenheimer, C., Gratuze, B., Boucetta, S., Sanabani, A., and al-Mosabi, A.: Obsidian sources in highland Yemen and their relevance to archaeological research in the Red Sea region, J. Archaeol. Sci., 37, 2332–2345, https://doi.org/10.1016/j.jas.2010.04.007, 2010.
Klie, W.: Ostracoda, Muschelkrebse, in: Die Tierwelt Deutschlands und der angrenzenden Meeresteile, edited by: Dahl, F., 34, 1–230, Gustav Fischer Verlag, Jena, 1938.
Krstic, N.: Rod Candona (Ostracoda) iz Kongerijskikh Slojeva Juzhnog dela Panonskog Basena (The genus Candona (Ostracoda) from Congeria Beds of the southern Pannonian Basin), Monographs of the Serbian Academy of Sciences and Arts section of Natural and Mathematical Sciences, 39, I–Vll, 1–145, 1972.
Lézine, A.-M., Saliège, J.-F., Robert, C., Wertz, F., and Inizan, M.-L.: Holocene Lakes from Ramlat as-Sab'atayn (Yemen) Illustrate the Impact of Monsoon Activity in Southern Arabia, Quaternary Res., 50, 290–299, https://doi.org/10.1006/qres.1998.1996, 1998.
Malz, H.: Heterocypris vel Cyprinotus? Ist die Morphologie des Gehäuses entscheidend für die Bestimmung rezenter Ostracoden-Gattungen?, Senckenbergiana lethaea, 57, 185–199, 1976.
Martens, K.: Ostracod speciation in ancient lakes, in: Speciation in ancient lakes, edited by: Martens, K., Goddeeris, B., and Coulter, G., Arch. Hydrobiol., 44, 203–222, 1994.
Mattash, M. A., Pinarelli, L., Vaselli, O., Minissale, A., Al-Kadasi, M., Shawki, M. N., and Tassi, F.: Continental Flood Basalts and Rifting: Geochemistry of Cenozoic Yemen Volcanic Province, International Journal of Geosciences, 4, 1459–1466, https://doi.org/10.4236/ijg.2013.410143, 2013.
Mayewski, P. A., Rohling, E. E., Stager, J. C., Karlen, W., Maasch, K. A., Meeker, L. D., Meyerson, E. A., Gasse, F., van Kreveld, S., and Holmgren, K.: Holocene climate variability, Quaternary Res., 62, 243–255, https://doi.org/10.1016/j.yqres.2004.07.001, 2004.
Mazzini, I. and Sardella, R.: Notes on the freshwater ostracods (Arthropoda: Crustacea) and on the Quaternary deposits of Socotra, Fauna of Arabia, 20, 181–191, 2004.
Mazzini, I., Anadon, P., Barbieri, M., Castorina, F., Ferreli, L., Gliozzi, E., Mola, M., and Vittori, E.: Late Quaternary sea-level changes along the Tyrrhenian coast near Orbetello (Tuscany, central Italy): palaeoenvironmental reconstruction using ostracods, Mar. Micropaleontol., 37, 289–311, 1999.
Mazzini, I., Gliozzi, E., Rossetti, G., and Pieri, V.: The Ilyocypris puzzle: A multidisciplinary approach to the study of phenotypic variability, Int. Rev. Hydrobiol., 99, 395–408, https://doi.org/10.1002/iroh.201301729, 2014.
McClure, H. A. and Swain, F. M.: Fresh-water and Brackish-water fossils. Quaternary Ostracoda from the Rub' al Khali (“Empty Quater”), Saudi Arabia, Actes du VI Colloque Africain de Micropaléontologie, Tunis 1974, Annales des Mines et de la Géologie, 28, 427–441, 1980.
McKenzie, K. G.: Illustrated generic key to South African continental Ostracoda, Annals of the South African Museum, 74, 45–103, 1977.
Meisch, C.: Freshwater Ostracoda of western and central Europe, in: Süsswasserfauna von Mitteleuropa, 8/3; Spektrum Akademischer Verlag Gustav Fischer, Heidelberg, Berlin, 2000.
Melovski, L., Matevski, V., and Hristovski, S.: Climate Change Vulnerability, Third national communication to UNFCCC, sector: biodiversity, climate change vulnerability (final report), Ministry of Environment and Physical Planning, Republic of Macedonia, 2013.
Mischke, S.: Mid and late Holocene palaeoenvironment of the lakes Eastern Juyanze and Sogo Nur in NW China, based on ostracod species assemblages and shell chemistry, Berliner Geowissenschaftliche Abhandlungen, Reihe E, 35, 1–131, 2001.
Mischke, S. and Almogi-Labin, A.: Quantitative reconstruction of lake conductivity in the Quaternary of the Near East (Israel) using ostracods, J. Paleolimnol., 43, 667–688, https://doi.org/10.1007/s10933-009-9359-y, 2010.
Mischke, S., Ginat, H., Al-Saqaratand, B., and Almogi-Labin, A.: Ostracods from water bodies in hyperarid Israel and Jordan as habitat and water chemistry indicators, Ecol. Indic., 14, 87–99, https://doi.org/10.1016/j.ecolind.2011.07.017, 2012.
Mischke, S., Ashkenazi, S., Almogi-Labin, A., and Goren-Inbar, N.: Ostracod evidence for the Acheulian environment of the ancient Hula Lake (Levant) during the early-mid Pleistocene transition, Palaeogeogr. Palaeocl., 412, 148–159, https://doi.org/10.1016/j.palaeo.2014.07.036, 2014.
Mischke, S., Opitz, S., Kalbe, J., Ginat, H., and Al-Saqarat, B.: Palaeoenvironmental inferences from late Quaternary sediments of the Al Jafr Basin, Jordan, Quaternary Int., 382, 154–167, https://doi.org/10.1016/j.quaint.2014.12.041, 2015.
Mohammed, M., Keyser, D., Al-Wosabi, M., Al-Khirbash, B., and Al-Qadassi, W.: Taxonomy and distribution of fresh water Ostracoda from Socotra Island, Yemen, Revue de Micropaleontologie, 57, 23–33, https://doi.org/10.1016/j.revmic.2013.10.001, 2013.
Moore, R. C.: Treatise on Invertebrate Paleontology, Part Q Arthropoda 3 Crustacea Ostracoda, Geological Society of America and University of Kansas Press, Lawrence, 1961.
Müller, G. W.: Deutschlands Süßwasser-Ostracoden, Zoologie, Original-Abhandlungen des Gesamtgebietes der Zoologie, 12, 1–112, 1900.
Özuluğ, O.: A preliminary study on Ostracoda (Crustacea) fauna of the Instranca streams, Turkey, Journal of Fisheries Sciences, 5, 93–98, https://doi.org/10.3153/jfscom.2011011, 2011.
Pachur, H. and Hoelzmann, P.: Paleoclimatic Implications of Late Quaternary Lacustrine Sediments in Western Nubia, Sudan, Qutaternary Res., 36, 257–276, 1991.
Paris, P.: Biospeologica 41: Ostracodes (Prem Ser), Archives Zoologie Experimentale, 58, 475–487, 1920.
Parker, A., Davies, C., and Wilkinson, T.: The early to mid-Holocene moist period in Arabia: some recent evidence from lacustrine sequences in eastern and south-western Arabia, Proc. Sem. Arab. Stud., 36, 243–255, 2006.
Ramdohr, F. A.: Über die Gattung Cypris Müll. und drei zu derselben gehörige neue Arten, Mag. Ges. Naturf. Berlin, 2, 83–93, 1808.
Rosenberg, T. M., Preusser, F., Fleitmann, D., Schwalb, A., Penkman, K., Schmid, T. W., Al-Shanti, M. A., Kadi, K., and Matter, A.: Humid periods in southern Arabia: Windows of opportunity for modern human dispersal, Geology, 39, 1115–1118, https://doi.org/10.1130/G32281.1, 2011.
Rosenfeld, A., Nathan, Y., Feible, C. S., Schilman, B., Halicz, L., Goren-Inbar, N., and Siman-Tov, R.: Palaeoenvironment of the Acheulian Gesher Benot Ya'aqov Pleistocene lacustrine strata, Northern Israel – lithology, ostracod assemblages and ostracod shell geochemistry, J. Afr. Earth Sci., 38, 169–181, https://doi.org/10.1016/j.jafrearsci.2003.10.008, 2004.
Sars, G. O.: Oversigt af Norges crustaceer med forelobige bemärkninger over de nye eller mindre bekjendte arter: 2 (Branchiopoda, Ostracoda, Cirripedia), Forhandlinger i Christiania Videnskabs-selskebet, 1890, 1–80, 1890.
Schöning, C.: Subrecent Ostracoda (Crustacea) from the Sudan, with a description of the juvenile stages of Oncocypris muelleri (DADAY, 1910), Mitteilungen des Hamburgischen Zoologischen Museum und Institut, 93, 39–56, 1996.
Sohn, I. G. and Morris, R. W.: Cheikella, a new fresh-water ostracode genus, and Telekia, a new name for a homonym, Micropaleontology, 9, 327–331, 1963.
van Harten, D.: Some new shell characters to diagnose the species of the Ilyocypris gibba-biplicata-bradyi group and their ecological significance, in: Proceedings of the VII International Symposium on Ostracodes, Taxonomy, Biostratigraphy and distribution of Ostracodes, edited by: Krstic, N., Serbian Geological Society, Belgrad 1979, 71–76, 1979.
Wilkinson, T. J.: Holocene environments of the high plateau Yemen: recent geoarchaeological investigations, Geoarchaeology, 12, 833–864, 1997.