the Creative Commons Attribution 4.0 License.
the Creative Commons Attribution 4.0 License.
 | 27 May 2024
| 27 May 2024
Rediscovering Globigerina bollii Cita and Premoli Silva 1960
Alessio Fabbrini
Maria Rose Petrizzo
Isabella Premoli Silva
Luca M. Foresi
Bridget S. Wade
Globigerina bollii Cita and Premoli Silva was described from the historical Langhian-type section in Langhe, Piedmont (Italy). Due to its peculiar compact morphology, it was set apart from all the other globigerinids typical of the coeval Mediterranean fauna, and it was only reported for a short and limited stratigraphic range. The taxon became a first-order marker for the local biostratigraphy with its own Globigerina bollii Zone within the Langhian stage. However, the species was later synonymised with Globigerina falconensis Blow, ending its use in biostratigraphic schemes, and it was no longer utilised by authors working in the Mediterranean area and Paratethys. We present a reassessment of Globigerina bollii, showing for the first time a full collection of high-quality scanning electron and optical microscope images of the type series of specimens and a comparative study with Mediterranean individuals from the Langhian of the Cretaccio Section (Italy) and extra-Mediterranean individuals from Ocean Drilling Program Site 747 in the Kerguelen Plateau (Indian Ocean). The stratigraphic ranges of all the occurrences cited in the scientific literature from 1960 to the present day and all the references including images of the taxon are compiled. We compare G. bollii to other four-chambered morphospecies inhabiting the oceans during the Miocene, providing a detailed discussion of their morphological differences, which allows us to retain G. bollii as a valid taxon and to disclaim its synonymy with Globigerina falconensis. Our taxonomical observations also allow us to reassign Globigerina bollii to the genus Globoturborotalita, due to its strong affinities with other members of that genus, such as G. eolabiacrassata Spezzaferri and Coxall, and G. ouachitaensis (Howe and Wallace). We present a direct visual comparison with the other representatives of middle Miocene globoturborotaliids. An additional comparison is also discussed with Globigerina bollii lentiana Rögl, a species endemic in the Paratethys. We conclude that the presence of G. bollii in the Mediterranean Basin during such a confined stratigraphic interval (Mediterranean Subzone MMi4c–MMi4d), might be a palaeogeographical indicator of the intermittent opening of the eastern gateway with the Paratethys, affecting the Mediterranean faunas during the Langhian and their migration from oceanic realms into the Paratethys and Mediterranean.
- Article
(7274 KB) - Full-text XML
- BibTeX
- EndNote





Several species of planktonic foraminifera characterised by four chambers in the final whorl have existed since the origin of the first planktonic foraminifera family, Conoglobigerinidae in the Late Jurassic (Petrizzo et al., 2020). This morphology appears to be widespread across the evolutionary history of planktonic foraminifera. Additional features determine variability at the genus and species level (for instance, lips and rims and apertural shape and height), but all these features still have an unknown role in the ecology or lifestyle of the organisms. These characteristics emerge during speciation events, separating subpopulations from each other and ultimately generating new species. For these reasons, the classification of planktonic foraminifera is challenging and faces several issues due to convergent evolution, where similar features evolve multiple times in the fossil record. Four chambers in the final whorl seems to be one of these recurrent and basic attributes.
After the Late Jurassic, foraminifera with four chambers in the final whorl never really disappeared, and they are still common today across several genera. Even after mass extinctions, such as at the K–Pg boundary, these basic four-chambered forms, e.g. Eoglobigerina, were among the first groups to reappear in the fossil record (Olsson et al., 2006; Morard et al., 2022). All of these similarities and convergent evolution have constituted a problem in the classification. Numerous authors have interpreted the morphological variability among populations with different levels of tolerance while describing morphospecies. Taxonomical atlases (Pearson et al., 2006; Wade et al., 2018) have been published to clarify decades of mismatching concepts and problematic morphospecies. Nevertheless, the Cenozoic family Globigerinidae possesses several species with a similar number of chambers in the last whorl (Fig. 1), averaging at four, and it still needs a deep revision, especially for the Neogene.
The limited morphological variability and high tendency to similarity creates a big problem not only for taxonomy but also for biostratigraphy and biochronology, challenging authors to identify morphospecies sometimes based on subjective and hardly detectable differences. Spezzaferri et al. (2015), Wade et al. (2018), Poole and Wade (2019), Fayolle and Wade (2021), Fabbrini et al. (2021, 2023), and Latas et al. (2023) showed how taxonomy can benefit from integrated methodologies based progressively more on statistics, biometry, and high-detail imaging, which were not available in the past. Here we address one of these problematic taxa, Globigerina bollii Cita and Premoli Silva 1960, a marker species in biostratigraphic schemes of the Mediterranean and Paratethys areas, which was then almost forgotten due to a questionable synonymy with Globigerina falconensis Blow, 1959 (Fig. 1). Our study contributes to new efforts to reassess the taxonomy and stratigraphic distributions of species by the Neogene and Quaternary Planktonic Foraminifera Working Group (NQPFWG), including taxonomic variability and the establishment of junior synonyms.

Figure 1Holotype comparison of (1) Globigerina falconensis Blow, 1959, and (2) Globigerina bollii Cita and Premoli Silva, 1960. The scale bar is 100 µm.
Taxonomic history
Blow (1959) described numerous four-chambered taxa, among which was a compact Globigerina characterised by a distinctive apertural lip, which he named Globigerina falconensis. This species was described from the lower Miocene of Venezuela at the topmost part of the Burdigalian. Subsequently, Globigerina falconensis was widely adopted and consequently reported globally, extending to the present (Fabbrini et al., 2023). Cita and Premoli Silva (1960) did not report Globigerina falconensis in their study of the Langhian stratotype planktonic foraminifera from the Bricco della Croce section in Italy (Fig. 1). Cita and Premoli Silva (1960) described a new species exclusive of the Langhian, and they named it Globigerina bollii after Hans Bolli. The taxon quickly became a well-established biostratigraphic marker for the Langhian across the Mediterranean Basin (Cita and Premoli Silva, 1960; Cita et al., 1965; Cita and Premoli, 1968; Cita and Blow, 1969) and remained valid until Crescenti (1966). Blow (1969) placed Globigerina bollii Cita and Premoli Silva, 1960, as a junior synonym of Globigerina falconensis Blow. The synonymy obscured the taxon, but some authors kept recognising G. bollii, not considering the synonymy with G. falconensis as valid. Foresi et al. (2001) disagreed on the synonymy due to some morphological differences, such as G. bollii being globose and bigger than G. falconensis, with more inflated chambers and fewer incised sutures. Foresi et al. (2001) added that G. bollii tends to be kummerform, and in cases where the last chamber is normal, the difference with G. falconensis is even more evident, due to the absence of the lip characterising G. falconensis. Both Foresi et al. (2001) and Di Stefano et al. (2008) reported and provided an updated taxonomical discussion of G. bollii and considered it to an auxiliary marker for the Langhian Mediterranean biostratigraphy.
Globigerina bollii was also reported by Rögl (1969) in the Paratethian realm. He described, from lower Miocene sediments, a new taxon, Globigerina bollii lentiana, which he considered ancestral to G. bollii. The stratigraphic range of G. bollii lentiana in the Paratethys was constrained to upper Burdigalian and lower Langhian by Rögl (1985). Globigerina bollii lentiana is possibly a junior synonym of Globoturborotalita ouachitaensis (Spezzaferri et al., 2018), but further investigations are still required. On the other hand, specimens from Szekely and Filipescu (2016) from Transylvanian sediments have been synonymised with Globoturborotalita eolabiacrassata in Spezzaferri et al. (2018), highlighting how this group of globoturborotaliids from the Paratethys is difficult to classify due to the poor local fossil preservation.
Fossil preservation in outcrops can vary significantly, and correct fossil identification can be challenging. In the past, authors used to describe fossil faunas and report the entire assemblage but only illustrated the marker species, as providing drawings was necessary when light or scanning electron microscope (SEM) images were not available. In certain cases, no images were provided or available, making every subsequent study difficult. Globigerina bollii has been reported and identified by multiple authors across the Mediterranean Basin, but the lack of images or drawings does not allow a direct control on the concept applied by the authors to identify Globigerina bollii. We present here a chronological review of all occurrences of Globigerina bollii in the scientific literature (Table 2).
We studied samples from Langhian records from three Mediterranean outcrops: Bricco della Croce, Cretaccio, and Moria sections, where Globigerina bollii was listed and imaged by previous authors (Cita and Premoli Silva, 1960; Foresi et al., 2001; Di Stefano et al., 2008). We imaged specimens and compared them with the type specimens from Cita and Premoli Silva (1960) to assess the consistency of the taxonomical concept applied in the literature. The type material included several specimens which were marked as topotypes, along with the paratypes and the holotype. We imaged, with the SEM, the paratype specimens and holotype collected by Cita and Premoli Silva (1960) which consisted of 19 specimens from Bricco della Croce section (Italy). We imaged the topotype specimens under a light microscope.
We examined 20 samples from Cretaccio Section (Italy), from sample TCRE47 to TCRE131 (Foresi et al., 2001); 10 extra samples from Bricco della Croce section (Italy); and 12 samples (MOR200–MOR322) from Moria section (Italy) (Di Stefano et al., 2008), but no images were taken due to very poor fossil preservation. To compare the Mediterranean assemblage to the oceanic realm, we picked specimens from the same interval from Ocean Drilling Program (ODP) Site 747 from the Kerguelen Plateau (southern Indian Ocean). We examined and imaged three samples from ODP Hole 747A, namely 8H4 10–12 cm, 11H5 10–12 cm, and 12H2 110.5–112.5 cm. Samples were dry-sieved at 125 µm. Taxonomic concepts and species identification were based on the literature (Cita and Premoli Silva, 1960; Bolli et al., 1985; Foresi et al., 2001; Spezzaferri et al., 2018). All the biostratigraphic data presented in this paper refer to the zonation of Wade et al. (2011) for the oceanic occurrences and to Lirer et al. (2019) for the Mediterranean occurrences and ranges.
2.1 Optical imaging
Selected specimens from Cretaccio Section (Italy) have been cleaned using an ultrasonic bath for 30–60 s at 50–60 Hz at room temperature (20 °C) in a plastic tube immersed in distilled water at the Department of Earth Sciences, University College London. The ultrasonic cleaning allowed the partial removal of infilling and encrustation on the tests to expose apertural features and wall texture details. The specimens were then transferred into microslides and imaged using a multifocal camera mounted on a stereo microscope at the Department of Earth Sciences, University College London.
2.2 Scanning electron microscope imaging
The specimens from Cretaccio Section (Italy) were placed on metal stubs using double-sided sticky tape. The stubs were coated with gold and imaged using a JEOL JSM-6480LV high-performance, variable pressure analytical scanning electron microscope (SEM) at the Department of Earth Sciences, University College London. The holotype and paratypes of Globigerina bollii were imaged at the Department of Earth Sciences, University of Milan, using a JSM-IT 500 (JEOL-2019) scanning electron microscope. The type specimens were imaged without gold coating in order to preserve their surface and appearance.
The holotype of Globigerina bollii shows poor fossil preservation, but some key features can still be noted, such as its strongly compact test and the kummerform last chamber equipped with a thin apertural lip. Our SEM and light microscope images (Figs. 2, 3, and 4) show how the holotype and paratypes possess four chambers in the final whorl, slowly increasing in size and forming a compact test. The type specimens are very consistent in terms of morphology and size; all specimens are large (> 300 µm), with the maximum diameter of the holotype being 400 µm.
Table 1Comparison of the key features distinguishing Globoturborotalita bollii (Cita and Premoli Silva, 1960) from similar taxa within the genus Globoturborotalita.
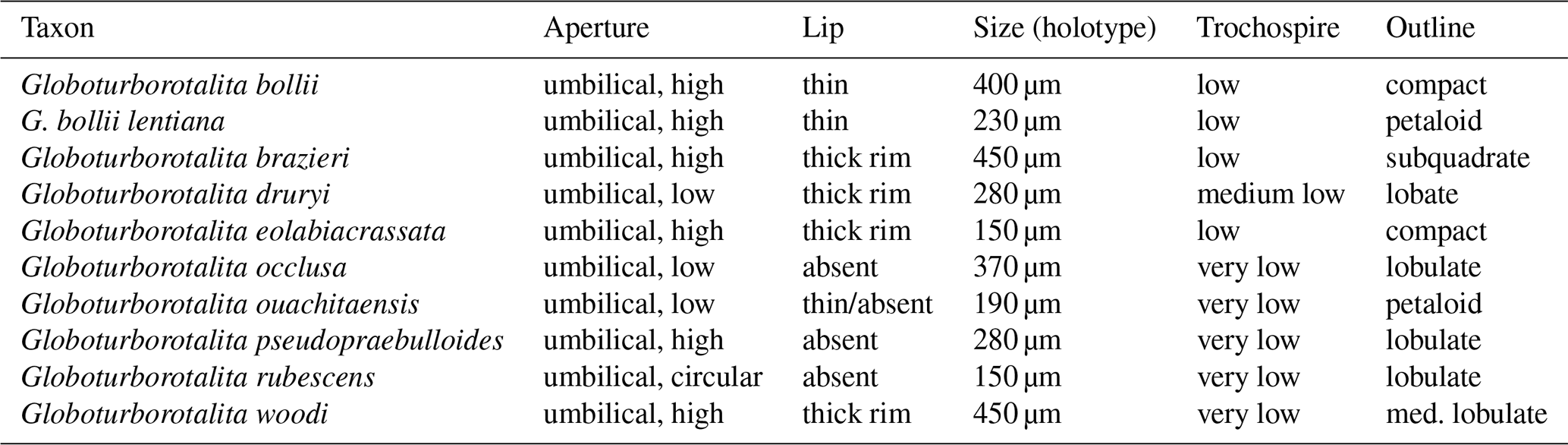
Table 2Occurrences of Globoturborotalita bollii and Globigerina bollii lentiana in the Mediterranean area, Paratethys, and oceanic realm. The LO (lowest occurrence) and HO (highest occurrence) of taxa in each section are indicated, adopting the Mediterranean biostratigraphic scheme from Lirer et al. (2019) and using information provided in the original publications.
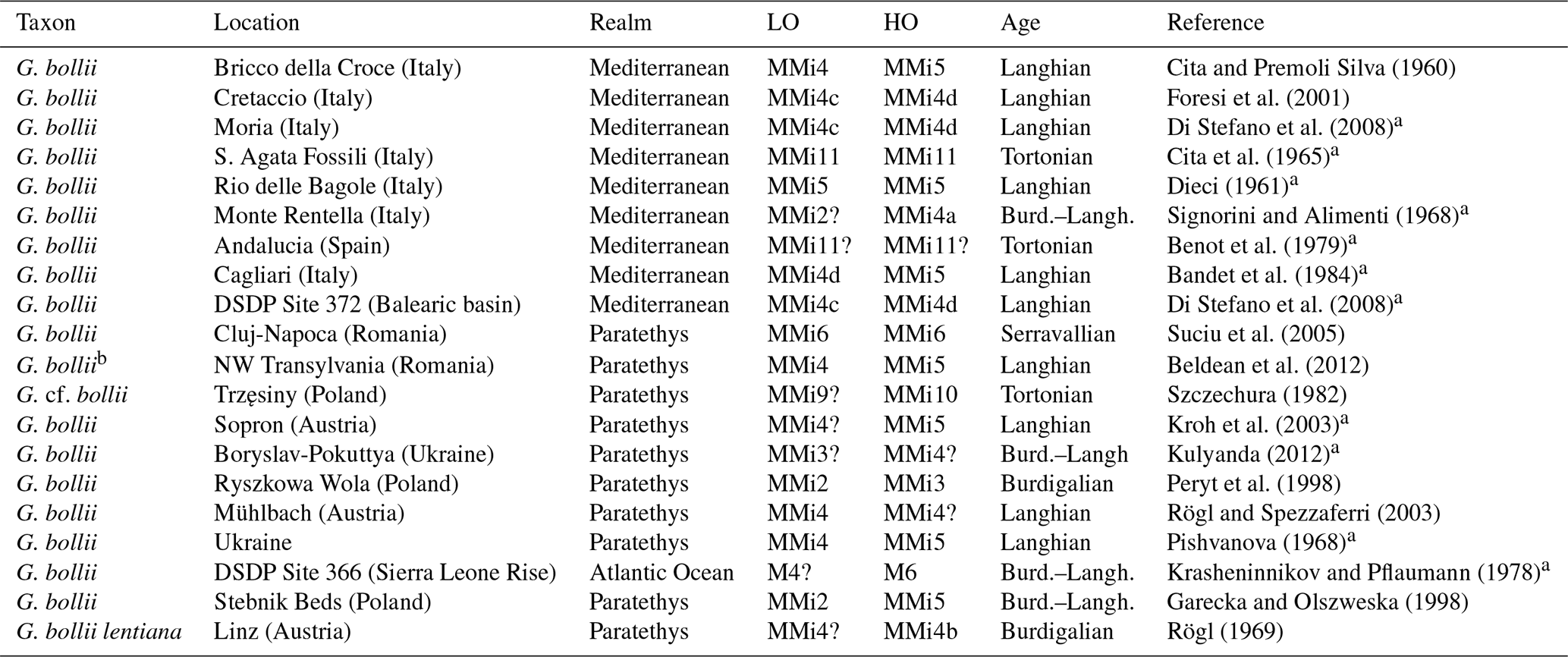
a Refers to articles not providing images or drawings of the taxa. b Named Globigerina gnaucki in the original paper.
The specimens picked at Cretaccio Section and at ODP Site 747 share the same morphological features and overall size. In both cases, the fossil preservation is generally good, with some infilling and signs of recrystallisation. Despite the infilling, the aperture, in these cases, is clearly visible and consists of a rounded central, low, and umbilical aperture that is normally bordered by a thin lip.
At ODP Site 747, previous work was conducted by Berggren (1992) and Verducci et al. (2009), where they recorded specimens attributed to Globoturborotalita brazieri and to Globoturborotalita woodi plexus (shown in Majewski, 2010), respectively. We examined specimens in our samples from ODP Site 747 and imaged and compared them to G. bollii (Fig. 5). Our specimens share strong similarities with G. bollii Cita and Premoli Silva, such as the compact outline, the tendency towards kummerform individuals, an arched aperture with a lip, and a comparable test size. This group of Globoturborotalita occurs between samples 12H2–8H4 in our study, equivalent to early-Miocene zones of M1–M2 (Wade et al., 2011).

Figure 2Holotype comparison in the umbilical view, with a new SEM of the holotype of Globigerina bollii Cita and Premoli Silva, 1960, Globigerina bollii lentiana Rögl, 1969, Globoturborotalita eolabiacrassata Spezzaferri and Coxall, 2018, and Globoturborotalita ouachitaensis Howe and Wallace, 1932. The scale bar is 100 µm.

Figure 3New SEMs of the holotype and paratypes of Globigerina bollii Cita and Premoli Silva. (1a–c) G. bollii Cita and Premoli Silva; holotype from collection number 1275 Bricco della Croce section (Langhe, Piedmont), Langhian, Subzone MMi4c. (2a–c) G. bollii paratype 1. (3a–c) G. bollii paratype 2. (4a–c) G. bollii paratype 3. (5a–c) G. bollii paratype 4. (6a–c) G. bollii paratype 5. (7a–c) G. bollii paratype 6. (8a–c) G. bollii paratype 7. (9a–c) G. bollii paratype 8. (10a–c) G. bollii paratype 9. Scale bar is 100 µm.
4.1 Taxonomic concept
The imaging we conducted on the type series material at the University of Milan allowed us to reexamine the overall taxonomical status of Globigerina bollii Cita and Premoli Silva. Our new light microscope and SEM investigations reveal an extremely consistent morphology at the multiple locations studied (Figs. 2–5). All specimens have a compact outline with a low trochospiral coiling resulting in a rhomboidal-subquadrate test, with an arched aperture bordered by a lip. Test sizes of G. bollii are consistently large (> 300 µm). Because of these attributes, we reject the synonymy, which placed G. bollii as junior synonym of G. falconensis. Globigerina falconensis shows a low trochospiral test arranged in about two whorls and with four subspherical chambers in the last whorl (Fig. 1). Globigerina falconensis also presents a small and deep umbilicus almost closed by the strongly developed lip on the last chamber (Fabbrini et al., 2023). “Globigerina” bollii can be distinguished from G. falconensis by the subquadrate shape, the less lobulate outline, and a more squared profile. For all of these reasons, the synonymy suggested by Crescenti (1966) and Blow (1969) should be rejected. We therefore consider G. bollii a valid taxon.

Figure 4SEMs of the type material used in the original paper from Cita and Premoli Silva (1960). (1a–c) G. bollii paratype 10 from sample 1279 Bricco della Croce, Langhian, Zone MMi4. (2a–c) G. bollii paratype 11. (3a–c) G. bollii paratype 12. (4a–c) G. bollii paratype 13. (5a–c) G. bollii paratype 14. The scale bar is 100 µm.
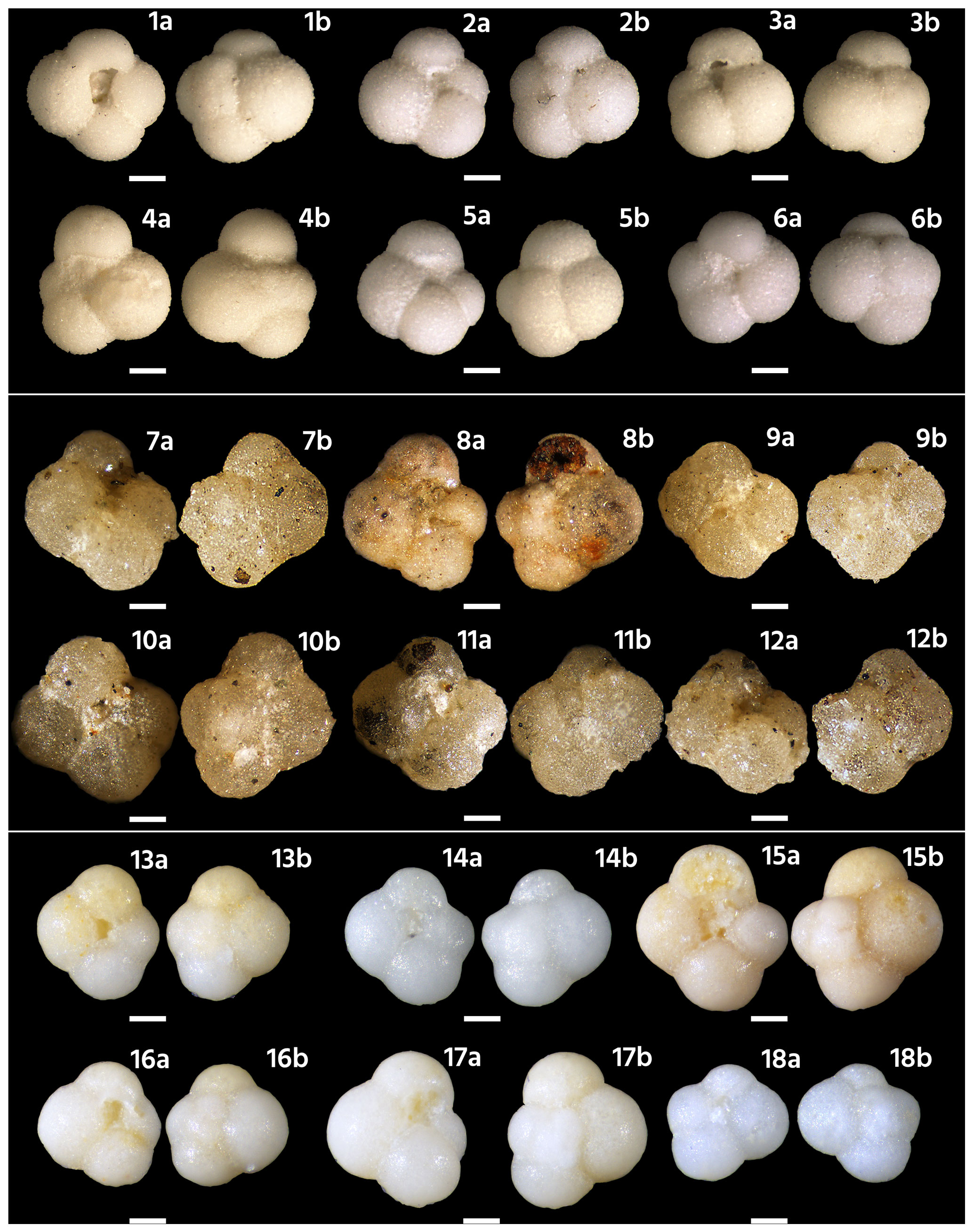
Figure 5Light microscope images of selected Langhian specimens of Globoturborotalita bollii. (1)–(6) Micro-Unimi nos. 2078, 2079, 2080, 2081, 2082, and 2083. ODP Hole 747A 8H4 10–12 cm, Kerguelen Plateau, Zone M7. (7)–(12) Globoturborotalita bollii Cita and Premoli Silva 1960 topotypes from sample 1260, Bricco della Croce section (Italy), Langhian, Subzone MMi4c (Zone N9 in original paper). (13)–(18) Micro-Unimi nos. 2084, 2085, 2086, 2087, 2088, and 2089. Globoturborotalita bollii from sample TCRE74, 26.80 m (Di Stefano et al., 2008), from Cretaccio Section (Italy), Subzone MMi4c. The scale bar is 100 µm.
Our new images of some of the paratypes suggest a cancellate wall texture ruber or sacculifer type, even though the fossil preservation of the holotype makes any definitive wall reassessment challenging. Nevertheless, G. bollii shows strong affinities with the genus Globoturborotalita rather than Globigerina when considering its compact test morphology and the thick wall appearance. Numerous morphospecies of Globoturborotalita occurred in the Miocene, all characterised by four chambers in the final whorl and a compact test. We compare G. bollii to these taxa to highlight differences and to support its taxonomical validity and its suggested reassignment to the genus Globoturborotalita. Therefore, we will refer to this taxon from now on as Globoturborotalita bollii.
Globoturborotalita bollii (Cita and Premoli Silva, 1960) has a distinctive morphology, which can be compared with other four-chambered taxa, inhabiting the oceans during the early to middle Miocene, such as Globoturborotalita eolabiacrassata Spezzaferri and Coxall, 2018 (in Spezzaferri et al., 2018), Globoturborotalita ouachitaensis (Howe and Wallace, 1932), and its putative ancestor Globigerina bollii lentiana Rögl, 1969 (Fig. 2). The holotype of Globoturborotalita bollii shows poor fossil preservation, but some key features can still be noted, such as its strongly compact test and the kummerform last chamber equipped with a thin apertural lip. Globoturborotalita ouachitaensis is a small taxon described from Eocene sediments in Ouachita, Louisiana (USA). This taxon has a distinctive petaloid outline, a loosely coiled test, and a very low to flat trochospire, with well separated chambers. Globoturborotalita ouachitaensis also presents a narrow umbilicus and a low straight aperture with a very thin lip. Globoturborotalita bollii is bigger than G. ouachitaensis, especially if comparing specimens from the Eocene and Oligocene (Spezzaferri et al., 2018), even though some specimens imaged from Subzone M1a (Aquitanian) shown in Spezzaferri et al. (2018) appear to be larger and more similar in size to the holotype of Globoturborotalita bollii. The maximum diameter measured on the holotype of G. bollii is about 400 µm, which makes this type bigger than the other taxa listed above (Table 1). Regardless of the size, the two morphospecies can be separated by the different test shape and lobate outline characterising G. ouachitaensis (Fig. 2).
Globoturborotalita eolabiacrassata (Fig. 2) was described from the early Miocene of the Kerguelen Plateau in Subzone M1a (Spezzaferri et al., 2018). This species has a small and compact test with maximum diameter of 150 µm, measured on the holotype, and it is characterised by a thick apertural rim. This species also presents a distinctive honeycomb cancellate wall texture when compared to both Globoturborotalita ouachitaensis (Howe and Wallace, 1932) and Globoturborotalita bollii (Cita and Premoli Silva, 1960). Globoturborotalita eolabiacrassata can be distinguished from G. bollii for its smaller size and the thick apertural rim.
The relation between Globoturborotalita bollii (Cita and Premoli Silva, 1960) and its alleged ancestor Globigerina bollii lentiana Rögl, 1969, is problematic because of their stratigraphic record inconsistencies and some morphological differences. In the original description, Rögl (1969) stated how G. bollii lentiana can be distinguished from G. bollii by the clearly separated chambers, the more incised sutures, the looser coiling, the deeper umbilicus, and the wider aperture. The final chamber is also usually significantly larger and also tilted over the depressed umbilical area in G. bollii lentiana and not in G. bollii. Globigerina bollii lentiana appears similar in its petaloid outline and small size to Globoturborotalita ouachitaensis. Spezzaferri et al. (2018) discussed how forms comparable to the holotype are rare, making a thorough comparison with G. bollii lentiana difficult. Some specimens previously identified as G. bollii lentiana have been synonymised with G. ouachitaensis, highlighting the morphological similarity between the two taxa. A reassessment of G. bollii lentiana will be addressed by the NQPFWG.
Globoturborotalita bollii shares some common features with G. eolabiacrassata and G. ouachitaensis, but it is distinct with regard to its subquadrate outline, apertural features (thin lip and high arched aperture), bigger size, and the limited stratigraphic distribution. Globoturborotalita bollii presents a thick and robust test (the wall texture verges towards a cancellate type similar to G. ouachitaensis), with a compact outline, four strongly embracing chambers in the last whorl, and incised intercameral sutures. All of these characteristics are not in line with the current concept of the genus Globigerina, which has bulloides-type wall texture.
Considering also the strong similarities between G. bollii, G. eolabiacrassata, G. ouachitaensis, and G. bollii lentiana, we can hypothesise that this group might share a common phylogeny. In this view, G. bollii lentiana and G. bollii might belong to the same lineage, evolving in the Paratethys and Mediterranean area during the Aquitanian–Burdigalian and Langhian, respectively. We provisionally suggest this phylogenetic relationship, but we keep all three morphospecies of G. ouachitaensis, G. bollii lentiana, and G. bollii separated due to the morphological differences discussed so far (Fig. 2 and Table 1). We also suggest reassigning G. bollii lentiana to the genus Globoturborotalita, pending a final assessment from the NQPFWG. A thorough taxonomical reassessment of this taxon and its phylogenetic relation with Globoturborotalita ouachitaensis is required. Following Rögl (1969), we provisionally retain G. bollii lentiana as the questionable ancestor of Globoturborotalita bollii.
4.2 Systematic palaeontology
-
Order Foraminiferida d'Orbigny, 1826
-
Superfamily Globigerinidae Carpenter, Parker and Jones, 1862
-
Family Globigerinidae Carpenter, Parker and Jones, 1862
-
Genus Globoturborotalita Hofker, 1976
-
Synonyms: Globigerina (Zeaglobigerina) Kennett and Srinivasan, 1983, p. 42
-
Type of species: Globigerina rubescens (Hofker, 1956)
-
Type of wall: Spinose, cancellate, and ruber- or sacculifer-type and sacculifer-type wall texture (Spezzaferri et al., 2018).
Test morphology. Test moderately low to medium trochospiral, globular, lobulate in outline and chambers globular; in a spiral view, there are three to five slightly embracing globular chambers increasing moderately in size in the last whorl; in the umbilical view, there are three to five globular, slightly embracing chambers, increasing moderately in size, with sutures depressed and straight on both sides. The umbilicus is generally small, open, and enclosed by surrounding chambers; in some species it can be larger. The aperture is umbilical, with a low to very high rounded arch, and bordered by a thin lip and rarely by a thickened rim. In the edge view, chambers are globular in shape, slightly embracing, and the umbilical aperture may be partly visible (modified after Spezzaferri et al., 2018, p. 234).
Remarks. Kennett and Srinivasan (1983) proposed Zeaglobigerina as a new subgenus accommodating Neogene Globigerina species with a cancellate wall, but Globoturborotalita Hofker 1976 is used, as it is a senior synonym. In the description of this genus, Hofker (1976) and Olsson et al. (2006) included only forms with four to five chambers in the last whorl. Spezzaferri et al. (2015) amended the genus to include also 3–3.5 chambered forms, such as G. connecta.
Range. Eocene (Zone E1) to recent (Spezzaferri et al., 2018).
-
Globoturborotalita bollii (Cita and Premoli Silva, 1960)
(Figs. 3–5).
-
1960 Globigerina bollii Cita and Premoli Silva, 1960, p. 120–125, fig. 1c, pl. XIII, figs. 1–18.
-
1967 Globigerina bollii Cita and Premoli Silva, 1960. Cita and Premoli Silva, p. 23, pl. 1, figs. 5a–c.
-
1969 ?Globigerina nilotica Viotti and Mansour, p. 443, pl. 6, figs. 5–8 (nomen dubium).
-
1970 ?Globigerina columbae Martinez Diaz, p. 1, Lámina 1, fig. 1c (nomen dubium).
-
1982 ?Globigerina cf. bollii Szczechura p. 40, figs. 8–15.
-
1992 ?Globoturborotalita brazieri Jenkins, 1965. Berggren, p. 644, pl. 1, fig. 4 [not Jenkins, 1965].
-
1998 Globigerina bollii Cita and Premoli Silva, 1960. Garecka and Olszewska, p. 717, pl. II, figs. 8a–8b.
-
1998 ?Globigerina bollii Cita and Premoli Silva, 1960. Peryt et al., p. 77-78, pl. 1, fig. 1, pl. 2, figs. 3–4.
-
2001 Globigerina bollii Cita and Premoli Silva, 1960. Foresi et al., p. 22, pl.1, figs. 22–24.
-
2003 Globigerina bollii Cita and Premoli Silva, 1960. Rögl and Spezzaferri, p. 10, pl. 10, fig. 15.
-
2005 Globigerina bollii Cita and Premoli Silva, 1960. Suciu et al., p. 458, pl. 1, fig. 7.
-
2012 Globigerina gnaucki Blow and Banner, 1962. Beldean et al., p. 183, pl. 3, figs. 7–8 [not Blow and Banner, 1962].
-
2012 ?Globigerina bollii Cita and Premoli Silva, 1960. Beldean et al., p. 184, pl. 4, fig. 7.
Type of wall. Spinose, cancellate, and ruber or sacculifer type?
Test morphology. Test compact and subquadrate, with a low trochospiral coiling mode and the profile poorly lobulate. The final whorl consists of four embracing spherical chambers, with the final chamber frequently smaller and slightly shifted toward the umbilicus. Straight sutures are weakly depressed and converging toward the umbilicus. Umbilical aperture is quite narrow and low bordered by a thin lip. Wall texture is difficult to determine on the holotype, with finer perforation on the last chamber. In the lateral and spiral view, the early whorls are hardly visible, thus the total number of chambers is not detectable.
Size. Holotype maximum diameter 400 µm (Figs. 1–3).
Remarks. The generic reattribution is based here on the general morphology of the test, the cancellate-like wall texture in some portion of the holotype (biased by the poor fossil preservation), and the numerous similarities between this taxon and Globoturborotalita ouachitaensis and G. eolabiacrassata. The assemblage recovered at Cretaccio Section (Italy) shows a robust wall texture not of the bulloides type and verging towards the ruber or sacculifer type.
Globoturborotalita bollii can be distinguished from Globigerina falconensis for having possibly a different wall texture and the subquadrate shape. Globigerina falconensis has a slightly more lobulate outline than G. bollii. The aperture in G. falconensis is a lower arch, which is more centred on the umbilicus and bordered by a thick lip. Moreover, Globoturborotalita bollii has a more squared profile and a thinner apertural lip than G. falconensis (see also Cita et al., 1965). For all of these reasons, the synonymy suggested by Crescenti (1966) and Blow (1969) is here rejected. Globoturborotalita bollii can be distinguished from Globigerina neofalconensis (Fabbrini et al., 2023) due to its squared outline and strongly compact test and higher asymmetrical aperture, which is in contrast to the lobulate outline and well-separated chambers and the well-developed apertural lip of G. neofalconensis.
We compare G. bollii to the other four-chambered Miocene globoturborotaliids (Table 1). It can be distinguished from G. eolabiacrassata by its bigger test size (Table 1), thinner apertural lip (rather than the thick rim in G. eolabiacrassata), and apparently a more finely perforated wall texture. It diverges from G. ouachitaensis by the overall bigger test (Table 1) and the more compact outline in contrast to the peculiar petaloid morphology of G. ouachitaensis. The aperture in G. bollii is higher and the umbilicus wider. It is separated from Globoturborotalita brazieri by the diamond shape of its test, and the number of chambers in the final whorl, which in G. brazieri tends to be 3–3.5 in the umbilical view. The rate of chamber growth is also more rapid in G. brazieri than in G. bollii, determining the overall different test morphology. A reverse droplet aperture characterises G. brazieri, while an asymmetrical low umbilical aperture is present in G. bollii. Globoturborotalita woodi (Jenkins, 1960) has a flatter spiral side, lower trochospire, more subquadrate morphology than G. bollii, and a higher symmetrical arched aperture without a lip.
Globoturborotalita bollii can be distinguished from G. pseudopraebulloides Olsson and Hemleben, 2018 (in Spezzaferri et al., 2018) by its more compact and subquadrate outline and the lower aperture bordered by a thin lip. The latter also displays a higher rate of chamber increase and fewer embracing chambers, giving this taxon a lobulate test. Globoturborotalita occlusa (Blow and Banner, 1962) differs from G. bollii with its lower, slit-like aperture without a lip and the extremely narrow umbilicus. Globoturborotalita rubescens (Hofker, 1956) differs from G. bollii in terms of its considerably smaller size (maximum diameter about 150 µm) and its circular high arched aperture. Globoturborotalita bollii differs from Globoturborotalita druryi (Akers, 1955) with respect to the well-developed and embracing final chamber, which is a more symmetrical umbilical aperture with a significantly thicker rim and tighter umbilicus.
Problematic taxa that are potentially related and share morphological similarities with Globoturborotalita bollii are Globigerina nilotica Viotti and Mansour, 1969, and Globigerina columbae Martinez Diaz, 1970, both described from the Mediterranean area. We consider these taxa nomina dubia that are waiting for future reexamination by the NQPFWG. Moreover, a definitive reconstruction of the phylogeny of G. bollii remains pending due to the limited specimens available and the generally poor fossil preservation of the Langhian assemblages in the Mediterranean and Paratethys.
Range. Mediterranean basin, from Subzone MMi4c to Subzone MMi4d (Lirer et al., 2019), within the magnetostratigraphic chrons C5Br–C5ADn (Lourens et al., 2004), and at ODP Site 747 in the zones O7–M5(?).
4.3 Globigerina bollii Zone in the Mediterranean
To determine the stratigraphic range of Globoturborotalita bollii in the Mediterranean, we review all of the reported occurrences and bring them together in an updated chronology (Fig. 6). Cita and Premoli Silva (1960) studied and described Bricco della Croce section (Italy) containing the historical stratotype of the Langhian stage (middle Miocene). Cita and Premoli Silva (1960) defined the new species G. bollii from the Cessole Formation at Bricco della Croce. The species here ranges from zones MMi4–MMi5 (Lirer et al., 2019) during the middle Langhian. Cita and Premoli Silva (1960) defined a biozone named the “Globigerina bollii Zone”, which falls into the Praeorbulina glomerosa Zone (Bolli, 1966) and Zone N8 (Blow, 1969). Following more recent zonal schemes, the range corresponds to the zones M5–M6 (Wade et al., 2011) and tentatively falls into Zone MMi4 (Lirer et al., 2019). The fossil preservation in this locality is poor, with recrystallised and heavily encrusted specimens.
Among the few inland sections with a good fossil preservation, the Cretaccio Section (Italy) (Fig. 6) is located on Cretaccio island in the Tremiti archipelago in southeastern Italy (Foresi et al., 1998, 2001; Di Stefano et al., 2008). At Cretaccio, G. bollii is reported as being abundant in the interval 0–40 m, which has been referred to as Zone MMi4. Foresi et al. (2001) documented, imaged, and discussed the presence of G. bollii from the base of the section (Langhian), ranging to below the first occurrence of Orbulina suturalis, within Subzone MMi4c (Lirer et al., 2019) and chron C5Bn.1r.
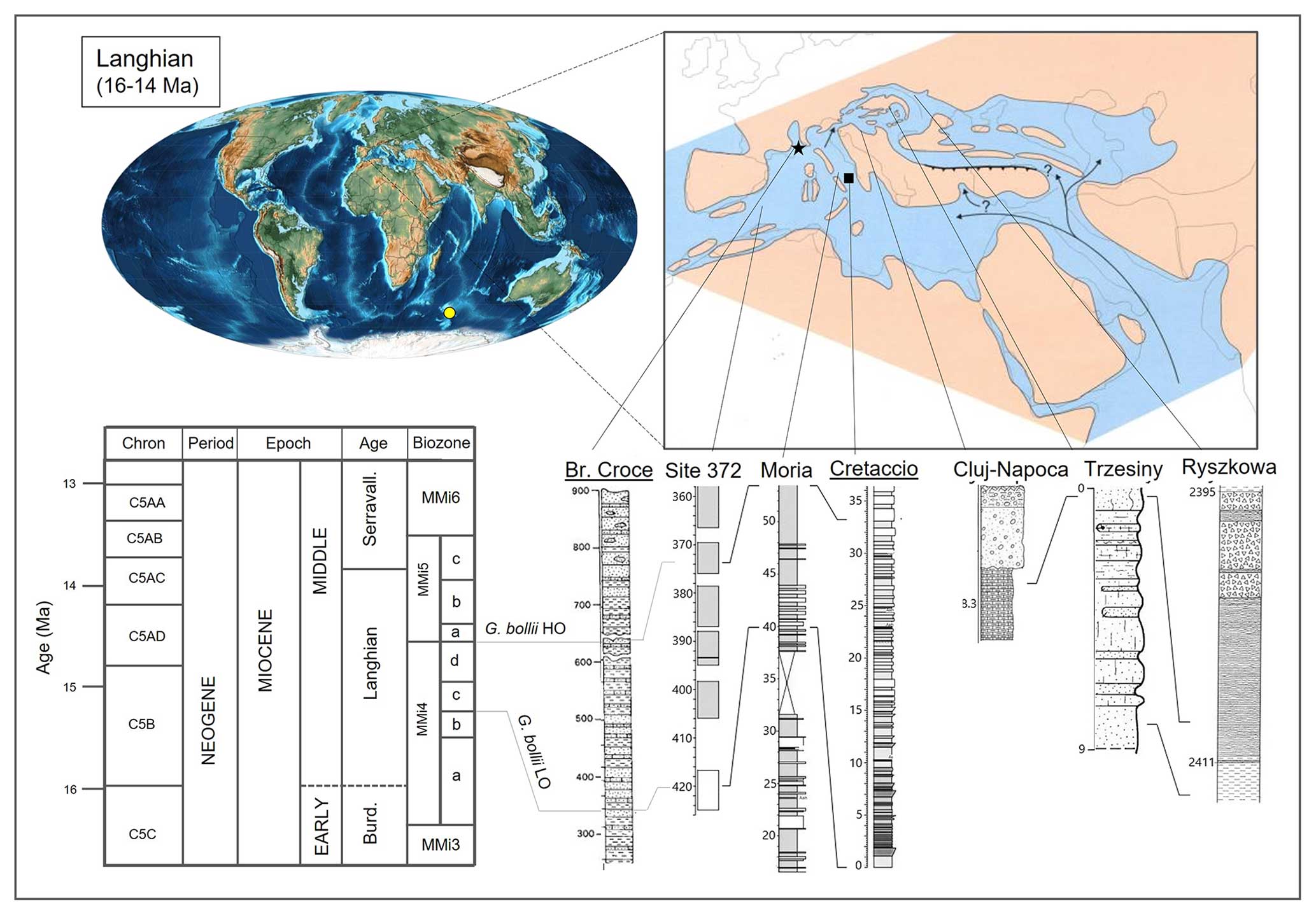
Figure 6Global palaeogeographic map during the Langhian (middle Miocene) modified after Scotese and Wright (2018). The right panel focuses on the palaeogeography in the Mediterranean Basin and Paratethys (modified after Rögl, 1998). The yellow dot shows the location of the extra Mediterranean record investigated at ODP Site 747 (Kerguelen Plateau). Selected stratigraphic sections where Globoturborotalita bollii was reported are indicated, and they are correlated to the Mediterranean zonation of Lirer et al. (2019). The star indicates the type of location of Globoturborotalita bollii (Cita and Premoli Silva, 1960) at Bricco della Croce (Italy). The reinvestigated outcrop (Cretaccio Section, Italy) is indicated with the black square. Globoturborotalita bollii LO (lowest occurrence) and HO (highest occurrence) are used for correlation between the Mediterranean area (Bricco della Croce, Deep Sea Drilling Project (DSDP) Site 372, Moria and Cretaccio), since the distribution range is well calibrated to magnetostratigraphic chrons C5Br to C5ADn and subzones MMi4c–MMi4d.
The Moria section is located in central Italy in the Marche region (Di Stefano et al., 2008). These sediments are rich in microfossils in the calcareous and marly portions. Di Stefano et al. (2008) reported G. bollii in a 14 m thick interval, correlated to Subzone MMi4c (chron C5Br), and with its HO (highest occurrence) in Subzone MMi4d (chron C5Bn.1r).
Globoturborotalita bollii was also cited in some other studies in the Mediterranean area with different degrees of confidence and information provided. Dieci (1961) reported the taxon in the Langhian Marne, a Pteropodi Formation in the Rio delle Bagole section, near Montegibbio, Modena, in the Apennines (Italy). Here G. bollii was described as abundant, and it has been used as a primary biostratigraphic marker for the Globigerina bollii Zone (Cita and Elter, 1960). These sandy marls can be dated to the Langhian, to Zone MMi5, due to the presence of O. suturalis and lying under the Dentoglobigerina altispira acme, representing Zone MMi6 (Serravallian stage) in Lirer et al. (2019).
A group of four-chambered taxa assigned to bulloides group was described in Cita et al. (1965), within the Tortonian stratotype in the Rio Mazzapiedi–Castellania section (S. Agata Fossili, Italy). In that section, Globoturborotalita bollii was reported as being rare, and for this reason, it was grouped together with other globigerinids. Cita et al. (1965) dedicated a paragraph to discuss Globoturborotalita bollii (Cita and Premoli Silva) and how this taxon could be distinguished from Globigerina falconensis, which is also present in the same assemblage. At Rio Mazzapiedi–Castellania, G. bollii ranges from the Globorotalia mayeri/Globorotalia lenguaensis Zone to the Globorotalia mayeri/Globoturborotalita nepenthes Zone (Blow, 1959). This interval can be correlated to Zone MMi11 (Tortonian–upper Miocene). Globoturborotalita bollii was reported by Signorini and Alimenti (1968) from Burdigalian and Langhian sediments in the Monte Penna area in central Italy. These sediments can be dated to Zone MMi2, due to the occurrence of C. dissimilis. Signorini and Alimenti (1968) also identified G. bollii within Marne di Corciano, a member of the Marnoso–Arenacea Formation, correlated to Subzone MMi4a.
Benot et al. (1979) reported and discussed G. bollii within Tortonian marls from Andalucia (Spain) within the Globorotalia acostaensis Zone and correlated to Zone MMi11 (Lirer et al., 2019) and tentatively to Zone M13 (Wade et al., 2011). Although the features listed by Benot et al. (1979) are consistent with the overall description of Globoturborotalita bollii, we are not able to determine the correct attribution of this taxon; thus this stratigraphic interval is listed in Table 2 of this paper, but it is still considered questionable for the total stratigraphic range of Globoturborotalita bollii in the Mediterranean area.
Another study on the Langhian stage in Sardinia, near Cagliari (Italy), conducted by Bandet et al. (1984), recorded G. bollii within Fangario clay Formation. These sediments can be correlated to Zone MMi5, due to the presence of both O. universa and O. suturalis. The authors dated biotites using KAr, obtaining a chronologic age of 15.30 ± 0.60 Ma, which corresponds to a Langhian age. No pictures or drawings were shown in the article.
The Deep Sea Drilling Project (DSDP) Site 372 was drilled in the eastern portion of the Menorca escarpment in the Mediterranean Balearic basin. Here Globoturborotalita bollii was documented from the bottom of the site (420 m b.s.f., below sea floor, in Core 31) up to 376 m b.s.f. in Core 27 (Bizon et al., 1978; Cita et al., 1978; Thunell, 1979; Di Stefano et al., 2008). This interval of blueish foraminifera–nannofossil marls are Langhian, and G. bollii was recognised in a confined range between Subzone MMi4c and MMi4d (Di Stefano et al., 2008).
4.4 Biochronology
Lirer et al. (2019) published an updated biostratigraphic scheme for the Mediterranean, with stratigraphic ranges of the main Neogene planktonic foraminifera and their local occurrences. Globoturborotalita bollii is included (named Globigerina bollii), and its stratigraphic range is between subzones MMi4c and MMi4d, with a question mark positioned at its base, highlighting the still-unknown level of appearance in the Mediterranean Basin, due to controversial mentions of the taxon below that level (Table 2). Foresi et al. (2001) is the primary source for Lirer et al. (2019) for the total range of Globoturborotalita bollii, which was reported in Cretaccio Section (Italy) as rare and only occurring in the Praeorbulina sicana glomerosa Subzone (Blow, 1959). The taxon predates the Praeorbulina glomerosa lowest occurrence (LO) in the Mediterranean basin (Di Stefano et al., 2008). The first typical specimens of G. bollii in Subzone MMi4c (Paragloborotalia siakensis–Praeorbulina circularis Subzone) were reported by Di Stefano et al. (2008) in all three sections (Moria, Cretaccio, and DSDP Site 372), occurring with Globorotalia praescitula, Catapsydrax parvulus, and Globorotalia archaeomenardii.
The occurrences in the Paratethys describe an older stratigraphic range for G. bollii, with some authors reporting its lowest local occurrence in the early Miocene within the Aquitanian/Burdigalian MMi2/MMi3 interval (Garecka and Oleszeska, 1998; Peryt et al., 1998; Kulyanda, 2012), thus predating the Mediterranean LO occurring in the Langhian (Zone MMi4). The occurrences from the Paratethys seem also to indicate a trend of progressively younger ranges (Zone MMi4–MMi5) in the Molasse Basin and in the central–western Paratethys, as shown at the Mühlbach section (Rögl and Spezzaferri, 2003) and at Sopron in modern Austria (Kroh et al., 2003), in the Transylvanian Basin (Beldean et al., 2012), and in Ukraine (Pishvanova, 1968). The regional differences make biostratigraphy correlations between the Mediterranean and Paratethys complex, adding a level of uncertainty in the stratigraphic distribution of G. bollii.
The only reported oceanic section mentioning G. bollii is located in the eastern equatorial Atlantic (DSDP Site 366), and it indicates also a LO older than the Mediterranean Basin, occurring within Zone MMi2 (Lirer et al., 2019) during the Burdigalian and is thus similar to what registered in the Paratethys. The absence of G. bollii in the Mediterranean Basin before the Langhian, but its occurrence during the Burdigalian in the Atlantic Ocean and in the Paratethys, might suggest that the species is not endemic in the Mediterranean area and that its evolution could be located somewhere else. Also, considering the open gateways present during the early Miocene between the Atlantic Ocean and the western Mediterranean Basin, the absence of G. bollii in DSDP Site 372 (Thunnell, 1979; Di Stefano et al., 2008) in levels lower than the Langhian can be an indication that the influx of this group of species into the Mediterranean might have occurred from the eastern gateway connecting the Mediterranean Basin with the Paratethys. This can be supported by the older stratigraphic range of the taxon in the Paratethys realm (tentatively MMi2–MMi5 interval). If older occurrences of G. bollii in the eastern Paratethys will be validated, then an original influx for G. bollii into the Paratethys from its eastern gateway (Indian Ocean) might be suggested.
Foresi et al. (2001) did not report the extinction of G. bollii. On the other hand, Ruggieri and Sprovieri (1970) put the extinction in Serravallian Zone N12 (Blow, 1969) (tentatively, Zone M10, sensu Wade et al., 2011), whereas Cita et al. (1965) and Cati and Borsetti (1968) identified the taxon ranging up to the Tortonian Zone N16 (Blow, 1969) and corresponding to Subzone M13a (Wade et al., 2011). The HO of Globoturborotalita bollii is reported within Zone MMi5 (Cita and Premoli Silva, 1968; Fornaciari et al., 1997; see also Foresi, 2001). The range at Site 366 spans to Zone M6 in the Langhian (Krasheninnikov and Pflaumann., 1978), with an age almost coeval to the HO of G. bollii elsewhere. According to Foresi et al. (2001) and Fornaciari et al. (1997), all occurrences above Zone N10 (Blow, 1969) (Zone M7, sensu Wade et al., 2011) would require reinvestigation to clarify a possible confusion between G. bollii and similar taxa.
4.5 Paratethys
Globoturborotalita bollii has also been reported outside of the Mediterranean area, occurring in several sections in the Paratethys basin with a different stratigraphic range (Table 2). The taxon was recognised in several outcrops, and G. bollii started to be used for the local biostratigraphy. Pishvanova (1968) and Pishvanova and Tkachenko (1971) used the Globigerina bollii Zone in Ukraine. Szczechura (1982) published a study on middle Miocene from Trzęsiny (Poland), reporting on the Badenian (middle Miocene) fossil assemblage with Globigerina cf. bollii. These specimens were recovered in association with Velapertina indigena, Trilobatus trilobus, orbulinids, Globigerina bulloides (?) (G. praebulloides in the original article) and nannofossils, and Bolboforma badenensis, which were correlated to the zones N13 and N14 (Blow, 1969), corresponding to zones M10–M11 (Wade et al., 2011).
Within the Carpathian Flysch in eastern Poland and western Ukraine, at the northern edge of the central Paratethys, Garecka and Olszweska (1998) reported Globoturborotalita bollii, showing a classic specimen in their plate 2, figs. 8a–b. The stratigraphic range of Globoturborotalita bollii is reported from early Burdigalian to late Langhian. Globoturborotalita bollii was also reported from the Badenian of Poland (Peryt et al., 1998).
Kulyanda (2012) recognised a Globoturborotalita bollii Zone within the local Karpatian age (upper Burdigalian) in Boryslav-Pokuttya (northwestern Ukraine). They reported no images but commented on the fossil assemblage characterising the local Karpatian stage, composed of Globoturborotalita bollii, Globigerina bulloides, Globigerina ottnangiensis, Trilobatus trilobus, Fohsella fohsi, and Globorotalia scitula. Such an assemblage can be correlated to the zonal interval MMi3–MMi4 (Burdigalian–Langhian).
Another occurrence of Globoturborotalita bollii was reported in Cluj-Napoca (Romania) from Serravallian sediments by Suciu et al. (2005), which can be correlated to Zone NN6 (Martini et al., 1971) using nannofossil markers, which falls into Zone MMi6 in Lirer et al. (2019). Still in Romania in the Transylvanian Basin during the Badenian age (i.e., Burdigalian/Langhian), both G. bollii and G. falconensis are documented by Beldean et al. (2012). Globigerina aff. bollii was mentioned in an outcrop located in Crimea during the Tarkhanian regional stage that was correlated to the Langhian stage (Popov et al., 2022). In that study, no images were provided; thus, any taxonomical assessment is not possible, but this recent paper shows how authors are still using the concept of this taxon. Globoturborotalita bollii and Globigerina falconensis were both reported and imaged in Mühlbach (Austria) within Gaindorf Formation in the Molasse Basin by Rögl and Spezzaferri (2003). The same interval also registered the presence of Praeorbulina glomerosa circularis transitional to Orbulina suturalis, which can indicate the Langhian Zone MMi5.
Rögl et al. (1969) described Globigerina bollii lentiana from Ottnangian sediments (corresponding to the middle Burdigalian stage and magnostratigraphic chrons C5E–C5D, following Piller et al., 2007), commenting on how G. bollii occurs in younger sediments (Langhian) in the Mediterranean.
4.6 Oceanic realm
The only mention in the literature of G. bollii outside the Mediterranean and Paratethian realm is from Krasheninnikov and Pflaumann (1978), where both G. bollii and G. falconensis are reported from the Sierra Leone Rise (DSDP Site 366) in the eastern equatorial Atlantic. The authors provided the local stratigraphic range of the taxon, but no images or drawings were given. Therefore, their taxonomic concept is not verifiable, and more examination is necessary to retain this record as valid.
Berggren (1992) recorded an abundant group of taxa described as Globoturborotalita brazieri and Globoturborotalita cf. labiacrassata at ODP Site 747. Both of these taxa do not correspond to Globoturborotalita brazieri, as shown in Spezzaferri et al. (2018), but instead they show strong morphological similarity with G. bollii. Verducci et al. (2009) reported high abundance of Globoturborotalita woodi at ODP Site 747, and this was also reported and imaged by Majewski (2010). In our opinion, these specimens are comparable with Globoturborotalita eolabiacrassata and to G. bollii. The specimens shown in Berggren (1992) are similar to the specimens we picked from the same interval in ODP Site 747 (Fig. 5). Berggren (1992) reported the total distribution of G. brazieri in zones O7 to M2 (Globoturborotalita brazieri Zone–Paragloborotalia incognita Zone in the original publication). We think that some of the taxa presented as G. brazieri in Berggren (1992) are consistent with G. bollii, thus potentially extending the fossil record of G. bollii to the uppermost Oligocene (Zone O7) outside of the Mediterranean. In this scenario, G. bollii would not be endemic in the Mediterranean basin but the result of immigration occurring in the Langhian.
4.7 Palaeogeographic observations
The short stratigraphic range of Globoturborotalita bollii in the Mediterranean Basin and Paratethys and of G. bollii lentiana in the Paratethys can also be discussed from another perspective. Both these taxa share strong similarities with Globoturborotalita ouachitaensis, G. eolabiacrassata, and their inferred ancestor G. bassriverensis, which had a wide geographical distribution and a long stratigraphic record from the Eocene to the Miocene. All these taxa are reported to inhabit the mixed layer of tropical oceans worldwide (Aze et al., 2011; Spezzaferri et al., 2018).
With the geographic distribution extended to ODP Site 747 (Fig. 5), in this context, G. bollii might be a cosmopolitan species whose abundance and occurrence may have been overlooked due to its affinity and similarities with other Globoturborotalita and often grouped together with G. woodi. Globoturborotalita bollii has a limited stratigraphic range during the Langhian in the Mediterranean Basin, which could be interpreted as a palaeogeographic indicator. The sudden appearance in the Mediterranean area of G. bollii might be an overlooked phenomenon, recording the changes occurring in the planktonic communities during the early–middle Miocene evolution of the Mediterranean due to its complex geodynamic framework. This time interval coincides with the Miocene Climatic Optimum (MCO), characterised by global warming and highest temperatures of the past 35 Myr and eustatic sea level changes (Miller et al., 2005). The Mediterranean Basin was affected by major palaeogeographic changes during the MCO (Cornacchia et al., 2021; Brandano et al., 2021, among others) due to regional geodynamic processes leading to the shoaling and intermittent closure of the Indian gateway (Rögl, 1999).
Piller et al. (2007) summarised the complex evolution of the connection between the central Paratethys and the Mediterranean area, resulting in the complete disconnection from the eastern Paratethys and then having a limited exchange of water masses during the early Miocene (Burdigalian). Piller et al. (2007) also pointed out how reopenings between the eastern Paratethys and the Indo-Pacific during the Langhian were likely. Considering the stratigraphic occurrences of Globoturborotalita bollii in the Mediterranean Basin (MMi4c–MMi4d zonal interval) during the Langhian, we can infer an influx of this group of Globoturborotalita in the Paratethys during the Burdigalian before the closure of its eastern gateway. Progressively, G. bollii might have migrated westwards in the central Paratethys, ultimately reaching the Mediterranean area during the Langhian. More detailed biostratigraphic data are necessary to verify this hypothesis. In this scenario, progressively younger occurrences of G. bollii from the eastern Paratethys to the Mediterranean would be expected. The scarcity of reliable geological sections and related palaeomagnetic age control and/or sufficient fossil preservation prevent such investigation.
The synonymy defined by Crescenti (1966) between Globigerina bollii Cita and Premoli Silva (1960) and Globigerina falconensis Blow (1959) influenced generations of workers to overlook Globigerina bollii, apart from few exceptions. A limited number of papers illustrated the specimens, making any taxonomic assessment difficult without restudying the samples. We presented new data and images of the type series of specimens of Globigerina bollii Cita and Premoli Silva (1960), which enabled us to reassess the taxonomic position of this taxon. The new images both from the SEM and the light microscope allowed us to reconsider the synonymy of Globigerina bollii with Globigerina falconensis and to suggest the validity of G. bollii as an independent morphospecies. This taxon shares morphological similarities with a group of Globoturborotalita, such as G. eolabiacrassata and G. ouachitaensis. The images of the type material and the collection of new specimens from Cretaccio Section (Italy) allowed us to reallocate G. bollii to the genus Globoturborotalita. Our observations suggest that Globoturborotalita bollii (Cita and Premoli Silva, 1960) still retains unique characteristics to distinguish it from the other taxa belonging to its genus and also from its inferred ancestor G. bollii lentiana.
The biostratigraphic comparison between all the occurrences reported in the literature shows how G. bollii has a limited range in the Mediterranean Basin confined to zonal interval MMi4c–MMi4d (Lirer et al., 2019). The occurrences of this species in the Paratethys are hard to verify due to the scarcity of high-quality images in the literature and the generally poor fossil preservation affecting Paratethian assemblages. The restricted range of G. bollii in the Mediterranean Basin could provide biostratigraphic value to the taxon as an auxiliary event for the Mediterranean area biochronology. The sudden appearance in the Mediterranean Basin of G. bollii might be a palaeogeographic signal of the reopening of the eastern gateway between the Paratethys and the Mediterranean Basin. In case this relation will be confirmed, the use of Globoturborotalita bollii would be a useful tool in palaeogeographic and biostratigraphic studies.
All of the imaged new specimens and the type specimens of Globoturborotalita bollii (Cita and Premoli Silva, 1960) are deposited at Department of Earth Sciences “A. Desio”, University of Milan, Milan (Italy).
AF and BSW conceptualised the research question. AF collected the micropalaeontological data, wrote the paper, and made the figures. MRP and IPS provided access to type specimens and analytical facilities. LMF provided access to samples from the Mediterranean sections and assisted with the correction of the paper. BSW corrected the draft and figures and provided analytical facilities and access to samples. All authors discussed the results and commented on the paper.
The contact author has declared that none of the authors has any competing interests.
Publisher’s note: Copernicus Publications remains neutral with regard to jurisdictional claims made in the text, published maps, institutional affiliations, or any other geographical representation in this paper. While Copernicus Publications makes every effort to include appropriate place names, the final responsibility lies with the authors.
The authors are grateful to Jim Davy (SEM Lab at UCL), and Stefania Crespi (SEM Lab at the University of Milan) for the technical support. The authors thank Silvia Spezzaferri for the helpful and insightful taxonomical discussions on the subject.
This research has been supported by the Natural Environment Research Council (grant no. NE/P019013/1) to Bridget S. Wade.
This paper was edited by Sev Kender and reviewed by Tracy Aze and one anonymous referee.
Akers, W. H.: Some planktonic foraminifera of the American Gulf Coast and suggested correlations with the Caribbean Tertiary, J. Paleontol., 29, 647–664, 1955.
Aze, T., Ezard, T. H., Purvis, A., Coxall, H. K., Stewart, D. R., Wade, B. S., and Pearson, P. N.: A phylogeny of Cenozoic macroperforate planktonic foraminifera from fossil data, Biol. Rev., 86, 900–927, 2011.
Bandet, Y., Burgois, J., Glacon, G., Gourinard, Y., Magne, J., and Muller, C.: Position du Langhien dans les echelles de chronologie biostratigraphique et geomagnetique, C. R. Acad. Sc. Paris, t. 299, Seire II, 10, 650–656, 1984.
Beldean, C., Filipescu, S., and Bălc, R.: Paleoenvironmental and biostratigraphic data for the Early Miocene of the north-western Transylvanian Basin based on planktonic foraminifera, Carpath. J. Earth Env., 7, 171–184, 2012.
Benot, C., Parra, A., and Diaz, Y. M. G.: Contirbucion mediante foraminiferos al studio de la problematica del Neogeno Andaluz, Publicaciones de la Universidad de Sevilla, Anales de la Universidad Hispalense Serie Ciencias, Vol. 22, 1979.
Berggren, W. A.: Neogene planktonic foraminifer magnetobiostratigraphy of the southern Kerguelen Plateau (Sites 747, 748, and 751), in: Proceedings Ocean Drilling Program, Sci. Res., 120, 631–647, 1992.
Bizon, G., Cita, M. B., Wright, R., and Mueller, C.: DSDP Leg 42A biostratigraphic range charts, DSDP Init. Repts., 42, 1095–1138, 1978.
Blow, W. H.: Age, correlation, and biostratigraphy of the upper Tocuyo (San Lorenzo) and Pozon Formations, eastern Falcon, Venezuela, Bull. Am. Paleontol., 39, 67–251, 1959.
Blow, W. H. and Banner, F. T.: The mid-Tertiary (Upper Eocene to Aquitanian) Globigerinaceae, in: Fundamentals of mid-Tertiary Stratigraphical Correlation, edited by: Eames, F. E., Banner, F. T., Blow, W. H., and Clarke, W. J., Cambridge University Press, Cambridge, 61–151, 1962.
Blow, W. H.: Late Middle Eocene to Recent planktonic foraminiferal biostratigraphy, in: Proceedings of the first international conference on planktonic microfossils (Leiden, EJ Brill.), 1, 199–422, 1969.
Bolli, H. M.: Zonation of Cretaceous to Pliocene marine sediments based on planktonic foraminifera, Asociación Venezolana de Geología, Minería y Petróleo, 9, 3–26, 1966.
Bolli, H. M. and Saunders, J. B.: Oligocene to Holocene low latitude planktic foraminifera, in: Plankton Stratigraphy, edited by: Bolli, H. M., Saunders, J. B., and Perch-Nielsen, K., Cambridge University Press, Cambridge, 155–262, 1985.
Brandano, M., Cornacchia, I., Catanzariti, R., and Tomassetti, L.: The Monterey Event in the Mediterranean platform to basin transition: The Guadagnolo Formation (Miocene, Prenestini Mountains, Central Apennines), Palaeogeogr. Palaeocl., 564, 110–177, 2021.
Carpenter, W. B.: Introduction to the Study of the Foraminifera: By William B. Carpenter, edited by: William, K. P. and Jones, T. R. Publ. Ray Soc., 22, 1862.
Cati, F. and Borsetti, A. M.: Notes on the evolution of some planktonic Foraminifera from the Miocene in Central Italy, Giornale di Geologia, 35, 253–262, 1968.
Cita, M. B. and Blow, W. H.: The biostratigraphy of the Langhian-Serravallian and Tortonian in the type sections in Italy, Riv. Ital. Paleont., 75, 549–603, 1969.
Cita, M. B. and Elter, G.: La posizione stratigrafica delle marne a Pteropodi delle Langhe della Collina di Torino ed il significato cronologico del Langhiano, Accad. Nazi. dei Lincei. Ser., 8, 360, 1960.
Cita, M. B. and Premoli Silva, I.: Globigerina bollii, nuova specie del Langhiano della Langhe, Riv. Ital. Paleontol. S., 66, 119–126, 1960.
Cita, M. B. and Premoli Silva, I.: Evolution of the planktonic foraminiferal assemblages in the stratigraphical interval between the type-Langhian and the type-Tortonian and biozonation of the Miocene of Piedmont, Giorn. Geol., 35, 1051–1082, 1968.
Cita, M. B., Premoli Silva, I., and Rossi, I.: Foraminiferi planktonici del Tortoniano-tipo, Riv. ital. Paleont. Stratigr., 66, 308–317, 1965.
Cita, M. B., Ryan, W. B., and Kidd, R. B.: Sedimentation rates in Neogene deep-sea sediments from the Mediterranean and geodynamic implications of their changes, Initial Rep. Deep Sea, 42, 991–1002, 1978.
Cornacchia, I., Brandano, M., and Agostini, S.: Miocene paleoceanographic evolution of the Mediterranean area and carbonate production changes: A review, Earth-Sci. Rev., 221, 103785, https://doi.org/10.1016/j.earscirev.2021.103785, 2021.
Crescenti U.: Sulla biostratigrafia del Miocene affiorante al confine marchigiano-abruzzese, Geol. Rom., 5, 1–54, 1966.
Dieci, G.: Pteropodi langhiani di Rio delle Bàgole (Montegibbio, Appennino settentrionale modenese), Boll. Soc. Paleont. Ital, 1, 31–42, 1961.
Di Stefano, A., Foresi, L. M., Lirer, F., Iaccarino, S. M., Turco, E., Amore, F. O., and Aziz, H. A.: Calcareous plankton high resolution biomagnetostratigraphy for the Langhian of the Mediterranean area, Riv. Ital. Paleontol. S., 114, 51–76, 2008.
d'Orbigny, A. D.: Tableau méthodique de la classe des Céphalopodes, in Annales des Sciences Naturelles, Series, 1, 96–314, 1826.
Fabbrini, A., Zaminga, I., Ezard, T. H., and Wade, B. S.: Systematic taxonomy of middle Miocene Sphaeroidinellopsis (planktonic foraminifera), J. Syst. Palaeontol., 19, 953–968, https://doi.org/10.1080/14772019.2021.1991500, 2021.
Fabbrini, A., Greco, M., Iacoviello, F., Kucera, M., Ezard, T., and Wade B. S.: Bridging the extant and fossil record of planktonic foraminifera: implications for the Globigerina lineage, Paleontology, 66, e12676, https://doi.org/10.1111/pala.12676, 2023.
Fayolle, F. and Wade, B. S.: The evolution of Eocene planktonic foraminifera Dentoglobigerina, J. Syst. Palaeontol., 19, 333–376, https://doi.org/10.1080/14772019.2021.1904021, 2021.
Foresi, L. M., Iaccarino, S., Mazzei, R., and Salvatorini, G.: New data on middle to late Miocene calcareous plankton biostratigraphy in the Mediterranean area, Rivista Italiana di Paleontologia e Stratigrafia, 104, 95–114, 1998.
Foresi, L. M., Iaccarino, S., Mazzei, R., Salvatorini, G., and Bambini, A. M.: Il plancton calcareo (foraminiferi e nannofossili) del Miocene delle Isole Tremiti, Paleontographia Italica, 88, 1–64, 2001.
Fornaciari, E., Iaccarino, S., Mazzei, R., Rio, D., Salvatorini, G., Bossio, A., and Monteforti, B.: Calcareous plankton biostratigraphy of the Langhian historical stratotype, in: Miocene Stratigraphy: An Integrated Approach. Developments in Palaeontology and Stratigraphy, 15, edited by: Montanari, A., Odin G. S., and Coccioni, R., Elsevier, Amsterdam, 89–96, 1997.
Garecka, M. and Olszewska, B.: Biostratigraphy of the Early Miocene of the Southern Poland based on planktic foraminifera and calcareous nannoplankton, Przegląd Geologiczny, 46, 712–721, 1998.
Hofker, J.: Foraminifera of Santa Cruz and Thatch-Island, Virginia-Archipelago West-Indies, Copenhagen Univ. Zool. Mus. Spolia (Skrifler), 15, 1–237, 1956.
Hofker, J.: La famille Turborotalitidae, Revue de Micropaléontologie, 19, 47–53, 1976.
Howe, H. V. and Wallace, W. F.: Foraminifera of the Jackson Eocene at Danville Landing on the Ouachita, Catahoula Parish, Louisiana (No. 13–15), Department of Conservation, Bureau of Scientific Research and Statistics, Minerals Section, 1932.
Jenkins, D. G.: Planktonic foraminifera from the Lakes Entrance oil shaft, Victoria, Australia, Micropaleontology, 6, 345–371, 1960.
Jenkins, D. G.: Planktonic foraminiferal zones and new taxa from the Danian to lower Miocene of New Zealand, New Zeal. J. Geol. Geop., 8, 1088–1126, 1965.
Kennett, J. P. and Srinivasan, M. S.: Neogene planktonic foraminifera, A phylogenetic atlas, Hutchinson Ross Pub. Co., 265 pp., 1983.
Krasheninnikov, V. A. and Pflaumann, U.: Zonal stratigraphy of Neogene deposits of the eastern part of the Atlantic Ocean by means of planktonic foraminifers, Leg 41 Deep Sea Drilling Project, in: Initial Reports of the Deep Sea Drilling Project, US Government Printing Office, 41, edited by: Lancelot, Y., Seibold, E., Dean, E. E., Jansa, L. F., Eremeev, V., Gardner, J., Cepek, P., Krasheninnikov, V. A., Pflaumann, U., Johnson, D., Rankin, J. G., and Trabant, P., Washington DC, 613–658, 1978.
Kroh, A., Harzhauser, M., Piller, W. E., and Rögl, F.: The Lower Badenian (Middle Miocene) Hartl Formation (Eisenstadt – Sopron Basin, Austria), in: Stratigrafia Austriaca, edited by: Piller, W. E., Österreichische Akademie der Wissenschaften, Schriftenreihe Erdwissenschaftliche Kommission 16, 87–109, 2003.
Kulyanda, M. and Hnylko, O.: Verification of the stratigraphy of Neogene molasses of the Boryslav-Pokuttya and Sambir Nappes in the north-west ern part of the Ukrainian Precarpathians, Heolohiyai Heokhimiya Horyuchykh Kopalyn, 158/159, 36–50, 2012.
Latas, M., Pearson, P. N., Poole, C. R., Fabbrini, A., and Wade, B. S.: Globigerinoides rublobatus – a new species of Pleistocene planktonic foraminifera, J. Micropalaeontol., 42, 57–81, https://doi.org/10.5194/jm-42-57-2023, 2023.
Lirer, F., Foresi, L. M., Iaccarino, S. M., Salvatorini, G., Turco, E., Cosentino, C., and Caruso, A.: Mediterranean Neogene planktonic foraminifer biozonation and biochronology, Earth-Sci. Rev., 196, 102869, https://doi.org/10.1016/j.earscirev.2019.05.013, 2019.
Lourens, L. J., Hilgen, F. J., Laskar, J., Shackleton, N. J., and Wilson, D.: The Neogene period, in: A geologic time scale 2004, edited by: Gradstein, F. M., Ogg, J. G., and Smith, A. G., Cambridge, Cambridge University Press, https://doi.org/10.1017/CBO9780511536045, 2004.
Majewski, W.: Planktonic foraminiferal response to middle Miocene cooling in the Southern Ocean (ODP Site 747, Kerguelen Plateau), Acta Palaeontol. Pol., 55, 541–560, https://doi.org/10.4202/app.2009.0088, 2010.
Martinez Diaz, C.: Tres nuevas especies de foraminiferos en el Andaluciense, Acts Geológica Hispánica, 5, 1–3, 1970.
Martini, E.: Standard Tertiary and Quaternary calcareous nannoplankton zonation, in: Proceedings Second Planktonic Conference, Rome, 739–785, 1971.
Miller, K. G., Kominz, M. A., Browning, J. V., Wright, J. D., Mountain, G. S., Katz, M. E., Sugarman, P. J., Cramer, B. S., Christie-Blick, N., and Pekar, S. F.: The Phanerozoic record of global sea-level change, Science, 310, 1293–1298, 2005.
Morard, R., Hassenrück, C., Greco, M., Fernandez-Guerra, A., Rigaud, S., Douady, C. J., and Kucera, M.: Renewal of planktonic foraminifera diversity after the Cretaceous Paleogene mass extinction by benthic colonizers, Nat. Commun., 13, 7135, https://doi.org/10.1038/s41467-022-34794-5, 2022.
Olsson, R. K., Hemleben, C., Huber, B. T., and Berggren, W. A.: Taxonomy, biostratigraphy, and phylogeny of Eocene Globigerina, Globoturborotalita, Subbotina, and Turborotalita, in: Atlas of Eocene Planktonic Foraminifera, Cushman Foundation of Foraminiferal Research, Special Publication, 41, edited by: Pearson, P. N., Olsson, R. K., Huber, B. T., Hemleben, C., and Berggren, W. A., 111–168, 2006.
Pearson, P. N., Olsson, R. K., Huber, B. T., Hemleben, C., and Berggren, W. A.: Atlas of Eocene Planktonic Foraminifera, Cushman Foundation of Foraminiferal Research, Special Publication, 41, 513 pp., 2006.
Peryt, T. M., Peryt, D., Szaran, J., Hałas, S., and Jasionowski, M.: O poziomie anhydrytowym badenu w utworze wiertniczym Ryszkowa Wola 7 k, Jarosławia (SE Polska), Biuletyn Państwowego Instytutu Geologicznego, 379, 61–78, ISSN 02086603, 1998.
Petrizzo, M. R., Wade, B. S., and Gradstein, F. M.: Evolution and biostratigraphy: Planktonic Foraminifera, in: Geologic Time Scale 2020, edited by: Gradstein, F. M., Ogg, J. G., Schmitz, M. D., and Ogg, G. M., 74–87, ISBN 9780128243602, 2020.
Piller, W. E., Harzhauser, M., and Mandic, O.: Miocene Central Paratethys stratigraphy–current status and future directions, Stratigraphy, 4, 151–168, 2007.
Pishvanova, L. S.: On the zonation of the Miocene by means of planktonic foraminifera, G. Geol., 35, 233–254, 1968.
Pishvanova, L. S. and Tkachenko, O. F.: Paleogeograficheskie karty miocena zapadnykh oblastei Ukr. SSR. Földtani Közlöny, Bull. Hung. Geol. Soc., 101, 265–276, 1971.
Poole, C. R. and Wade, B. S.: Systematic taxonomy of the Trilobatus sacculifer plexus and descendant Globigerinoidesella fistulosa (planktonic foraminifera), J. Syst. Palaeontol., 17, 1989–2030, https://doi.org/10.1080/14772019.2019.1578831, 2019.
Popov, S. V., Golovina, L. A., Palcu, D. V., Goncharova, I. A., Pinchuk, T. N., Rostovtseva, Y. V., Akhmetiev, M. A., Aleksandrova, G. N., Zaporozhets, N. I., Bannikov, A. F., and Bylinskaya, M. E.: Neogene Regional Scale of the Eastern Paratethys, Stratigraphy and Paleontological Basis, Paleontol. J., 56, 1557–1720, https://doi.org/10.1134/S0031030122120024, 2022.
Rögl, F.: Die Foraminiferenfauna aus den Phosphoritsanden von Plesching bei Linz (Oberosterreich) – Ottnangien (Untermiozan), Sonderdruck aus Mitteilungen der Geologischen Gessellschaft in Wien, 213–234, 1969.
Rögl, F.: Late Oligocene and Miocene planktic foraminifera of the Central Paratethys, in: Plankton Stratigraphy, edited by: Bolli, H. M., Saunders, J. B., and Perch-Nielsen, K., Cambridge University Press, Cambridge, 315–328, 1985.
Rögl, F.: Palaeogeographic considerations for Mediterranean and Paratethys seaways (Oligocene to Miocene), Annalen des Naturhistorischen Museums in Wien, Serie A für Mineralogie und Petrographie, Geologie und Paläontologie, Anthropologie und Prähistorie, 279–310, 1998.
Rögl, F.: Mediterranean and Paratethys. Facts and hypotheses of an Oligocene to Miocene paleogeography (short overview), Geol. Carpath., 50, 339–349, 1999.
Rögl, F. and Spezzaferri, S.: Foraminiferal paleoecology and biostratigraphy of the Mühlbach section (Gaindorf Formation, lower Badenian), Lower Austria. Annalen des Naturhistorischen Museums in Wien, Serie A für Mineralogie und Petrographie, Geologie und Paläontologie, Anthropologie und Prähistorie, 23–75, 2003.
Ruggieri, G. and Sprovieri, R.: I microforaminiferi delle Marne di S. Cipirello, Lavori Ist. Geol. Palermo, 10, 1–26, 1970.
Scotese, C. R. and Wright, N.: PALEOMAP paleodigital elevation models (PaleoDEMS) for the Phanerozoic, PALEOMAP Project, 2018.
Signorini, R. and Alimenti, M.: La serie stratigrafica del Monte Rentella fra il Lago Trasimeno e Perugia, GEOL. ROM., VII, 75–94, 1968.
Spezzaferri, S., Kucera, M., Pearson, P. N., Wade, B. S., Rappo, S., Poole, C. R., and Stalder, C.: Fossil and genetic evidence for the polyphyletic nature of the planktonic foraminifera “Globigerinoides”, and description of the new genus Trilobatus, PLoS One, 10, e0128108, https://doi.org/10.1371/journal.pone.0128108, 2015.
Spezzaferri, S., Olsson, R. K., Hemleben, C., Wade, B. S., and Coxall, H. K.: Taxonomy, biostratigraphy, and phylogeny of Oligocene and lower Miocene Globoturborotalita, in: Atlas of Oligocene planktonic foraminifera, edited by: Wade, B. S., Olsson, R. K., Pearson, P. N., Huber, B. T., and Berggren, W. A., Cushman Foundation Foraminiferal Research, Special Publication, 46, 231–268, 2018.
Suciu, A. A., Chira, C., and Popa, M. V.: Late Badenian Foraminifera, calcareous nannofossils and pteropod assemblages identified in boreholes from Cluj-Napoca, Acta Palaeontol. Rom., 5, 451–461, 2005.
Székely, S. F. and Filipescu, S.: Biostratigraphy and paleoenvironments of the Late Oligocene in the north-western Transylvanian Basin revealed by the foraminifera assemblages, Palaeogeogr. Palaeocl., 449, 484–509, 2016.
Szczechura, J.: Middle Miocene foraminiferal biochronology and ecology of SE Poland, Acta Palaeontol. Pol., 27, 1982.
Thunell, R. C.: Mediterranean Neogene planktonic foraminiferal biostratigraphy: quantitative results from DSDP Sites 125, 132 and 372, Micropaleontology, 412–437, 1979.
Verducci, M., Foresi, L. M., Scott, G. H., Sprovieri, M., Lirer, F., and Pelosi, N.: The Middle Miocene climatic transition in the Southern Ocean: evidence of paleoclimatic and hydrographic changes at Kerguelen plateau from planktonic foraminifers and stable isotopes, Palaeogeogr. Palaeocl., 280, 371–386, 2009.
Viotti, C. and Mansour, A.: Tertiary planktonic foraminiferal zonation from the Nile Delta, U.A.R., Part II. Globigerina nilotica, a new species of foraminifera from the Miocene of the Nile Delta, in: Proceedings of the Third African Micropaleontological Colloquium, edited by: Said, R., Beckmann, J. P., Ghorab, M. A., El Ansary, S., Viotti, C., and Kerdany, M. T., Cairo March 1968, National Information and Documentation Centre, Cairo, 442–447, 1969.
Wade, B. S., Pearson, P. N., Berggren, W. A., and Pälike, H.: Review and revision of Cenozoic tropical planktonic foraminiferal biostratigraphy and calibration to the geomagnetic polarity and astronomical time scale, Earth-Sci. Rev., 104, 111–142, 2011.
Wade, B. S., Olsson, R. K., Pearson, P. N., Huber, B. T., and Berggren, W. A.: Atlas of Oligocene planktonic foraminifera, Cushman Foundation Foraminiferal Research, Special Publication, 46, 528 pp., 2018