the Creative Commons Attribution 4.0 License.
the Creative Commons Attribution 4.0 License.
 | 03 May 2024
| 03 May 2024
Progress in the taxonomy of Late Cretaceous high-latitude radiolarians: insights from the Horton River area, Northwest Territories, Canada
Juan F. Diaz
Noritoshi Suzuki
Jennifer M. Galloway
Manuel Bringué
Radiolarians have proven to be exceptional tools in biostratigraphic and paleoenvironmental studies despite their relatively low preservation potential in the sedimentary record. Reports of Upper Cretaceous radiolarians in high northern latitudes are scant, and their taxonomy is not often discussed in the few reports available in the literature. In this study, we describe one new genus and three new species of polycystine radiolarians from Upper Cretaceous strata of the northern mainland coast of Arctic Canada. The specimens were recovered from the Santonian–middle Campanian Smoking Hills Formation and the middle Campanian–Maastrichtian Mason River Formation and are assigned to the genus Canadasphaera n. gen. (Order Spumellaria) and the species Canadasphaera inuita n. sp., Kuppelella rusti n. sp., and Lithocampe? rara n. sp. The taxonomic notes provided in this study improve our understanding of Late Cretaceous radiolarians from high northern latitudes and serve as a foundation for future Cretaceous radiolarian research in Arctic regions.
-
Canadasphaera: urn:lsid:zoobank.org:act:C6B7B702-4B74-472D-8893-0A41B93A5066, date of registration: 19 April 2024
-
Canadasphaera inuita: urn:lsid:zoobank.org:act:C30E555B-EE94-4680-87D1-5CE83E128461, date of registration: 19 April 2024
-
Kuppelella rusti: urn:lsid:zoobank.org:act:68ECB628-2E74-4748-ACA4-DD1A33B05837, date of registration: 19 April 2024
-
Lithocampe? rara: urn:lsid:zoobank.org:act:25E569B2-93A8-425F-BB7C-D29A0437763F, date of registration: 19 April 2024
- Article
(5997 KB) - Full-text XML
- BibTeX
- EndNote





Upper Cretaceous radiolarian assemblages from high northern latitudes are scarce, low in diversity and generally poorly preserved compared to Upper Cretaceous records from other localities worldwide. This paucity of information has hindered efforts to create radiolarian biostratigraphic zonations in this part of the world and limited their use as paleoenvironmental proxies in Cretaceous strata. In the Arctic basins of North America, Upper Cretaceous radiolarians have only been reported from the Arctic Slope of northern Alaska (Tappan, 1960; Bergquist, 1966) and Ellef Ringnes Island in the Sverdrup Basin of Arctic Canada (Pugh et al., 2014). In the interior of North America, impoverished radiolarian assemblages have been recovered from the Pierre Shale Formation in the central United States (Schultz et al., 1980; Bergstresser, 1983) and Manitoba, Canada (Rust, 1892; Tyrrell, 1892; Young and Moore, 1994; Muehlbauer, 2014); the Lea Park and Bearpaw formations in Alberta, Canada (Wall, 1975); and the Niobrara Formation in southern Saskatchewan, Canada (Diaz and Velez, 2018). These studies provide information on the presence and vertical distribution of radiolarian species but do not address their taxonomy, which is crucial to constructing radiolarian biozones.
In this study, we describe one new genus and three new species of radiolarians recovered from the Smoking Hills and Mason River formations on the northern mainland coast of Arctic Canada (Figs. 1, 2). These units were deposited in a continental shelf environment (Plauchut and Jutard, 1976; Dixon, 1992) and are dated as Santonian–Maastrichtian based on fossil vertebrates (Russell, 1967), dinocysts (McIntyre, 1974), diatoms (Tapia and Harwood, 2002) and silicoflagellates (McCartney et al., 2011). The new radiolarian taxa described in this study include the genus Canadasphaera n. gen. (Order Spumellaria) and the species Canadasphaera inuita n. sp., Kuppelella rusti n. sp., and Lithocampe? rara n. sp. (Fig. 2). Diaz et al. (2020) provided a preliminary list of the radiolarian assemblage recovered from the Smoking Hills Formation. The full radiolarian assemblages from both units and their biostratigraphic significance will be discussed in a future contribution.

Figure 1Location map of the three sections analyzed in this study (Northwest Territories, Canada). Section 1 (red dot) and section 2 (blue dot) are located at 69°27′52′′ N, 126°58′13′′ W and 69°28′46′′ N, 126°59′20′′ W, respectively. Section 3 (green dot) is located at 69°58′19′′ N, 127°07′15′′ W. The radiolarians described in this study were recovered from sections 1 and 2. Section 3 is barren of radiolarians.
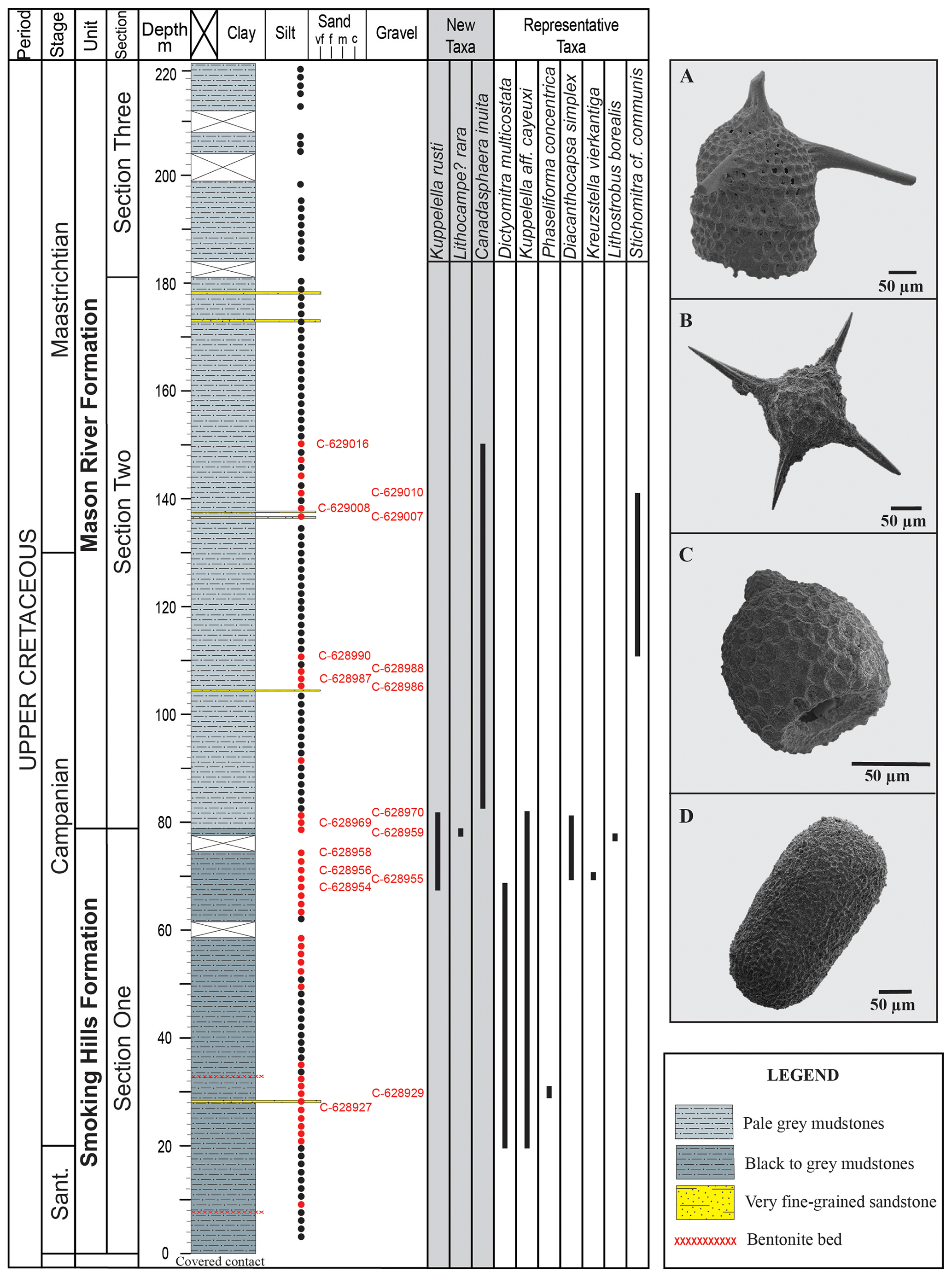
Figure 2Lithostratigraphy of the Smoking Hills and Mason River formations and stratigraphic distribution of the three new species described in this study and other representative radiolarian taxa recovered from both units. (A) Lithostrobus borealis Kozlova & Vishnevskaya in Vishnevskaya and Kozlova, 2012 (sample C-628959). (B) Kreuzstella vierkantiga Empson-Morin, 1981 (sample C-628956). (C) Kuppelella aff. cayeuxi (Squinabol, 1903) (sample C-628927). (D) Phaseliforma concentrica (Lipman, 1952) (sample C-628929). Black dots represent samples barren of radiolarians, and red dots represent samples where radiolarians were recovered. Red text indicates samples where the new and illustrated representative radiolarian taxa were identified.
2.1 The Smoking Hills Formation
The Smoking Hills Formation was defined by Yorath et al. (1975) at the type section along the Horton River in the Northwest Territories, Canada (Fig. 1, section 1). This section consists of 79 m of black to medium grey, soft but commonly fissile, bituminous mudstone (mainly in the lower part of the succession) with numerous jarosite bands that occur mainly in the upper part (Grasby et al., 2022). The presence of palynomorphs from Division H1 and H2 (McIntyre, 1974), diatoms from the Costopixis antiqua Zone (Tapia and Harwood, 2002), silicoflagellates from the Cornua trifurcata Zone (McCartney et al., 2011), and agglutinated foraminifera from the Glaphyrammina spirocompressa Zone (McNeil, 1997) in the Smoking Hills Formation indicate a Santonian–Campanian age for this unit. The lowermost 20 m of the Smoking Hills Formation may be as old as Coniacian, but the absence of all microfossil groups in this interval, except for a few palynomorphs (McIntyre, 1974), prevents a more accurate age determination. The Smoking Hills Formation is several hundred metres thick in the subsurface, as measured in wells on the Tuktoyaktuk Peninsula, ∼ 100 km to the west of the study area, and in parts of the southern Beaufort-Mackenzie Basin (e.g., Myhr, 1975; Dixon, 1992). At the type section, the Smoking Hills Formation rests disconformably on the Albian Horton River Formation, whereas in the subsurface it rests unconformably on Paleozoic to Turonian strata (Dixon, 1992).
2.2 The Mason River Formation
The Mason River Formation was defined by Yorath et al. (1975) at the type section along the Horton River in the Northwest Territories, Canada (Fig. 1, sections 2 and 3). This composite section consists of 143.5 m of pale to medium grey, blocky to fissile mudstone and sporadic very fine-grained sand layers. Ferruginous lithic fragments dominate the uppermost part of the unit. The contact between the Smoking Hills and Mason River formations is defined by an abrupt lithological colour change from black to pale grey. McNeil (1997) and Dixon and McNeil (2008) assigned an early Maastrichtian age to the Mason River Formation in the Beaufort-Mackenzie Basin based on the interpretation of dinoflagellate cyst assemblages of the Division H3 (McIntyre, 1974) and their analysis of benthic foraminifera. A late Campanian–early Maastrichtian age was proposed for the Mason River Formation by Tapia and Harwood (2002) and McCartney et al. (2011) based on diatoms (Trinacria indefinita and Stephanopyxis simonseni zones) and silicoflagellates (Arctyocha mesocena, Arctyocha quadralta and Lyramula furcula zones), respectively.
Although the Smoking Hills and Mason River formations exhibit similar lithology, their microfossil content is markedly different (Diaz et al., 2020). The Smoking Hills Formation is dominated by radiolarians with sporadic, agglutinated foraminifera, diatoms and silicoflagellates. In contrast, the Mason River Formation is dominated by diatoms and silicoflagellates, contains only a few radiolarian taxa and is barren of agglutinated foraminifera (Diaz et al., 2020). The Smoking Hills and Mason River formations were deposited in an outer shelf-to-slope environment of the epicontinental marine Anderson Basin (Yorath and Cook, 1981; Dixon, 1992), located at approximately 80° N in the convergence between the Western Interior and Sverdrup basins.
3.1 Field collection
Lithostratigraphic sections of the Smoking Hills and Mason River formations were described, measured and sampled (Smith et al., 2018; Diaz et al., 2020). The samples for this study were taken from the type sections of the Smoking Hills and Mason River formations described by Yorath et al. (1975) (Fig. 1). The type section of the Smoking Hills Formation is located at 69°27′52′′ N, 126°58′13′′ W. The basal conglomerate that marks the base of the Smoking Hills Formation, as described by Yorath et al. (1975), was not observed in the type section or any other areas during sample collection in 2018 (Smith et al., 2018). Dixon (1999) re-interpreted the basal conglomerate to be recent river terrace deposits juxtaposed against Smoking Hills strata. The Mason River Formation outcrops located at approximately 69°28′46′′ N, 126°59′20′′ W form a composite section that comprises the lower and middle parts of the formation. The third section is located at 69°58′19′′ N, 127°07′15′′ W and covers the upper part of the Mason River Formation (this section is barren of radiolarians, and it is not included in this study).
A total of 136 mudrock samples were collected from the sections for micropaleontological analyses. Samples were collected at regular increments of ∼ 1.5 m vertically throughout the Smoking Hills and Mason River formations. No macrofossils were observed. At each sample point, the surface material, largely unconsolidated mudstone, was removed, and unweathered material was sampled and placed in plastic bags and sealed. Thickness measurements and lithology are reported in Diaz et al. (2020) and Grasby et al. (2022).
3.2 Laboratory processing
Radiolarian shells were retrieved from sediment subsamples using the methods of Kennedy and Coe (2014) and Diaz and Velez (2017, 2018). Briefly, aliquots of 100 g were crushed, immersed in distilled water for 24 h, and repeatedly frozen and thawed until nearly all the sample was disaggregated. The remaining sediment was washed with household bleach, heated on an oscillating hot plate for 1 h and filtered through a 63 µm brass sieve. The dried residues were analyzed under a binocular microscope (WILD Heerbrugg) at 250× magnification. Classification of radiolarians was refined using a Quanta FEG 250 SEM and a Phenom Pro 6 Desktop SEM housed at the Department of Earth, Energy and Environment, University of Calgary. Some specimens (paratypes) were dissected with a razor blade to analyze their internal structures. This practice, although laborious, allowed for a more detailed description and accurate identification.
Classification at the superfamily and family levels follows that of Ehrenberg (1846), with subsequent emendations by De Wever et al. (2001) and Suzuki et al. (2021). Systematics at the species level includes a synonym list, derivation of name, diagnosis, holotype, material, range, description, dimensions, remarks and illustrations. Measurements were made on pictures using the software programs ImageJ and Adobe Illustrator. All measurements are given in micrometres (µm). The specimens described in this study are held in the National Collection of Invertebrate and Plant Fossil Types in Ottawa, Ontario, Canada.
-
Infrakingdom Rhizaria Cavalier-Smith, 2002 sensu emend. Cavalier-Smith (2003)
-
Phylum Retaria Cavalier-Smith 1999 stat. Cavalier-Smith (2002)
-
Subphylum Ectoria Cavalier-Smith in Cavalier-Smith et al. (2018)
-
Infraphylum Radiolaria Müller, 1859 sensu Adl et al. (2019)
-
Class Polycystinea Ehrenberg, 1839, stat. Levine et al. (1980)
-
Order Spumellaria Ehrenberg, 1876, stat. Haeckel 1884
-
Superfamily ? Haliommoidea Ehrenberg, 1846
-
Family ? Haliommidae Ehrenberg, 1846 sensu emend. Suzuki in Suzuki et al. (2021)
-
Genus Canadasphaera n. gen. Diaz et Suzuki
Type species. Canadasphaera inuita n. sp. Diaz et Suzuki (monotypy).
Derivation of name. Geographic name Canada and Greek sphaera, meaning “spheric”.
Diagnosis. Two spherical concentric shells without a microsphere. Distorted spherical outer concentric shell. Non-bladed radial spines except for their base near the cortical shell.
Description. Two spherical concentric shells without a microsphere. The cortical shell is distorted spherical, latticed, smooth on the surface, moderate in size and moderate in thickness. Several radial spines are visible on the cortical shell. They are thin, rod-like in shape and non-bladed except for their base near the cortical shell. The length of radial spines in one specimen may be variable. The medullary shell is spherical to hemispherical with a polygonal pore frame. Many radial beams connect the medullary shell to the cortical shell. These beams are distorted circular to distorted triangular in section, with wide distal ends, and arise from several triple junctions on the medullary shell.
Occurrence. Common in the lower and middle Mason River Formation (late Campanian–Maastrichtian).
Remarks. This new genus is marked by two spherical concentric shells; non-bladed radial beams that are distorted circular to distorted triangular in section; several non-bladed, rod-shaped radial spines; and a distorted spherical cortical shell. This genus does not include those with a perfectly spherical outermost concentric shell. The genus Canadasphaera is tentatively assigned to the Family Haliommidae (Superfamily Haliommoidea) because the double medullary shell characteristic of this family and superfamily was not observed in any specimens analyzed in this study. According to the integrated molecular and morphological classification, the morphology of radial beams and spines (bladed vs. non-bladed) is relevant to the family- and genus-level in the Haliommoidea (Suzuki et al., 2021). No precisely fitted type species to this new genus's character are recognized in any Cenozoic genera. No similar genera with a distorted spherical shape and non-bladed radial spines have been formally described in the literature as far as we know. Therefore, a new genus is established herein.
-
Canadasphaera inuita n. sp. Diaz et Suzuki
(Plate 1, figs. A–I) -
Cromyomma? nodosa Pessagno, 1976 Zonova et al., 1993, pl. XVIII, fig. 1.
Derivation of name. Named after Inuit (meaning “people” in Inuktitut) and the inhabitants of Arctic regions in Canada, Alaska and Greenland.
Diagnosis. Two spherical concentric shells without a microsphere. Distorted spherical cortical shell. Non-bladed radial spines except for their base near the cortical shell.
Holotype. Plate 1, fig. A, sample C-628990, deposited in the National Collection of Invertebrate and Plant Fossil Types in Ottawa, Canada, under repository number GSC-143273.
Material. 77 specimens recovered from 9 samples (C-628970, C-628986, C-628987, C-628988, C-628990, C-629007, C-629008, C-629010, and C-629016) in the lower and middle Mason River Formation (Fig. 2).
Range. Late Campanian–Maastrichtian.
Description. The cortical shell is distorted spherical and 2.13 to 2.39 times larger than the medullary shell. Pores on the cortical shell are relatively moderate in size. The pore frame is polygonal, but the pore shape is circular to elliptical. The pore frame is weakly rimmed on the outer surface. The triple junction has no distinctive accessories except for the radial spine. The exact number of radial spines is uncertain, but more than 7 and less than 12. These radial spines are relatively irregularly arranged on the cortical shell and never form a geometric arrangement. No direct connection of radial spine to radial beam is observed. Radial spines are thin rod-like in shape and non-bladed except for their base near the cortical shell. The length of radial spines in one specimen may be variable (between 62.5 and 69.1 µm). The tip of the radial spines is acute. The medullary shell is spherical to hemispherical. The pore frame of the medullary shell is thinner than that of the cortical shell. The pore frame is smooth on its surface. Several radial beams radiate from the medullary shell to connect with the cortical shell. Radial beams arise from the ratio of 40 % triple junctions on the medullary shell ( triple junctions) as far as known.
Dimensions (average based on nine specimens). Diameter of medullary shell: 46 µm; diameter of cortical shell: 106 µm; diameter of pores on the cortical shell: 10.1 µm; length of radial spine: 63.5 µm; number of pores on the half hemisphere of the cortical shell: 11; size ratio of the cortical shell with medullary shell: 2.26; size ratio of pores on the cortical shell with the diameter of the cortical shell: 0.10; size ratio of radial spine to the outer cortical shell: 0.68.
Remarks. Canadasphaera inuita morphologically resembles Acanthosphaera? sp. (Basov and Vishnevskaya, 1991, pl. 12, fig. 9) and Cromyomma? nodosa Pessagno, 1976 (Vishnevskaya, 1988, pl. II, fig. 6; Zonova et al., 1993, p. 42, pl. XVI, fig. 1). However, these two species have thin and robust three-bladed spines, respectively. A specimen classified as Cromyomma? nodosa Pessagno, 1976 (Zonova et al., 1993, p. 42, pl. XVIII, fig. 1) shows non-bladed spines, and it is here included as a synonym of Canadasphaera inuita.

Plate 1(A–I) Canadasphaera inuita n. sp. (A holotype number GSC-143273). (A–D) Sample C-628990: (A–C) view of the cortical shell, (D) view of the medullary shell. (E) Sample C-629007: view of the cortical shell. (F) Sample C-629010: view of the medullary shell. (G) Sample C-628990: view of the cortical and medullary shells. (H) Sample C-629007: view of the medullary shell. (I) Sample C-628990: view of the medullary shell.
-
Order Nassellaria Ehrenberg, 1876
-
Superfamily Pterocorythoidea Haeckel 1882 stat Kozur and Mostler 1984 sensu Suzuki in Suzuki et al., 2021
-
Family ?Lophocyrtiidae Sanfilippo and Caulet in De Wever et al., 2001
-
Genus Kuppelella Empson-Morin, 1981
-
Dictyocephalus Ehrenberg, 1861 (Squinabol, 1903, p. 132, pl. 8, fig. 28).
-
Tricolocapsa (Tricolocapsium) Haeckel, 1882 (Campbell and Clark, 1944, p. 35).
Type species. Tricolocapsa (Tricolocapsium) granti Campbell and Clark, 1944.
Description. Test commonly with two or three segments. Very large hemispherical thorax relative to very small cephalis. Cephalis spherical, simple. Relict pores scattered on the cephalis. No apical horn. Thorax hemispherical to rounded cylindrical. Abdomen, when present, variable with species but generally large, inflated and perforate (Empson-Morin, 1981). Aperture is always present, large and possesses an apertural ring on its mouth. No feet.
Remarks. Nassellaria having a simple poreless cephalis with no apical horn and lacking true feet belong to the superfamily Eucyrtidioidea and Pterocorythoidea (Sandin et al., 2021; Suzuki et al., 2021). The combination of a small cephalis and large robust thorax is characterized in Lophocyrtiidae Sanfilippo and Caulet in De Wever et al. (2001). We tentatively place this genus to the Lophocyrtiidae. Some species fitting the description of the genus Kuppelella Empson-Morin, 1981 have been previously assigned to the genus Dictyocephalus Ehrenberg, 1861 (e.g., Dictyocephalus microstoma, Rust, 1892, pp. 105–106, pl. XIV, fig. 5; Dictyocephalus cayeuxi, Squinabol, 1903, p. 132, pl. VIII, fig. 28; Dictyocephalus (?) lepidosus Kozlova in Kozlova and Gorbovetz, 1966, p. 103, pl. V, fig. 3; Dictyocephalus annikae Garg and Jain, 1978, pp. 157–158, pl. 4, fig. 59, pl. 5, fig. 80, pl. 6, fig. 99). However, the use of this genus name involves two serious problems. First, the genus name was already preoccupied by a senior homonym, Dictyocephalus Leidy 1856 [type species: Dictyocephalus elegans Leidy 1856]. Second, the type species to this genus is now identified as Dictyocryphalus obtusus (Ehrenberg, 1844), which belongs to the family Dictyocryphalidae in the superfamily Plagiacanthoidea (O'Dogherty et al., 2021; Suzuki et al., 2021). Schadelfusslerus Empson-Morin 1981 [type species: Schadelfusslerus echtus Empson-Morin 1981] is similar to Kuppelella, but the former always has distinctive three feet. Myllocercion Foreman 1968 [type species: Myllocercion acineton Foreman 1968] is fundamentally different from Kuppelella and Schadelfusslerus by having carpocaniid-type cephalis (Suzuki et al., 2021).
Included species. Kuppelella sp. aff. K. cayeuxi (Squinabol, 1903) [syn. Dictyocephalus (?) lepidosus Kozlova in Kozlova and Gorbovetz, 1966]; Dictyocephalus annikae Garg and Jain, 1978; Kuppelella sp. aff. K. granti (Campbell and Clark, 1944); Kuppelella rusti n. sp. Diaz et Suzuki.
-
Kuppelella rusti n. sp. Diaz et Suzuki
(Plate 2, figs. A–G)
Derivation of name. Named after David Rüst, the first author to describe and illustrate some species of Cretaceous radiolarians in North America.
Diagnosis. Small test with two or three segments. Very large hemispherical thorax relative to very small spherical cephalis. No apical horn and feet. Circular pores with spine-like protuberances at vertices. Large aperture and smooth circular rim.
Holotype. Plate 2, figs. A–B, sample C-628959, deposited in the National Collection of Invertebrate and Plant Fossil Types in Ottawa, Canada, under repository number GSC-143277.
Material. 209 specimens recovered from 7 samples (C-628954, C-628955, C-628956, C-628958, C-628959, C-628969 and C-628970) in the uppermost part of the Smoking Hills Formation and lowermost part of the Mason River Formation (Fig. 2).
Range. Late Campanian.
Description. Small test with two or three segments. Sub-spherical cephalis and dome-shaped to hemispherical thorax. The cephalis is small in absolute size, poreless, with a slight relict pore trace. No apical horn. Externally, the collar stricture is pronounced with a marked change in contour. Internally, the collar plate is pierced by four adjacent collar pores, each with a slightly different size. Thorax is large (about 3 to 5 times the size of the cephalis) and perforate. Pores are circular to subcircular, surrounded by polygonal pore frames with spine-like protuberances at vertices, giving an irregular texture to the test. Aperture is large, simple and has a smooth circular rim. Some specimens show evidence of a third chamber (abdomen), but it is always broken or not fully developed in the samples analyzed here.
Dimensions (average based on 12 specimens). Total height of test: 80 µm (n=12); width of cephalis: 25 µm (n=12); height of thorax: 75 µm (n=12); maximum width of thorax: 100 µm (n=12); diameter of aperture: 47 µm (n=5); diameter of the polygonal rimmed pore frame: 10 µm (n=5); number of pores in the half hemisphere of thorax: 12; number of pores in a vertical line of thorax: 7; width ratio of cephalis to thorax: 2.5; size ratio of pore to the maximum width of thorax: 0.10.
Remarks. This species differs from Kuppelella sp. aff. K. cayeuxi (Squinabol, 1903) and other species assigned to the genus Kuppelella, in having a more irregular pore pattern on the thorax, a smaller cephalis-to-thorax ratio, and a generally smaller size.
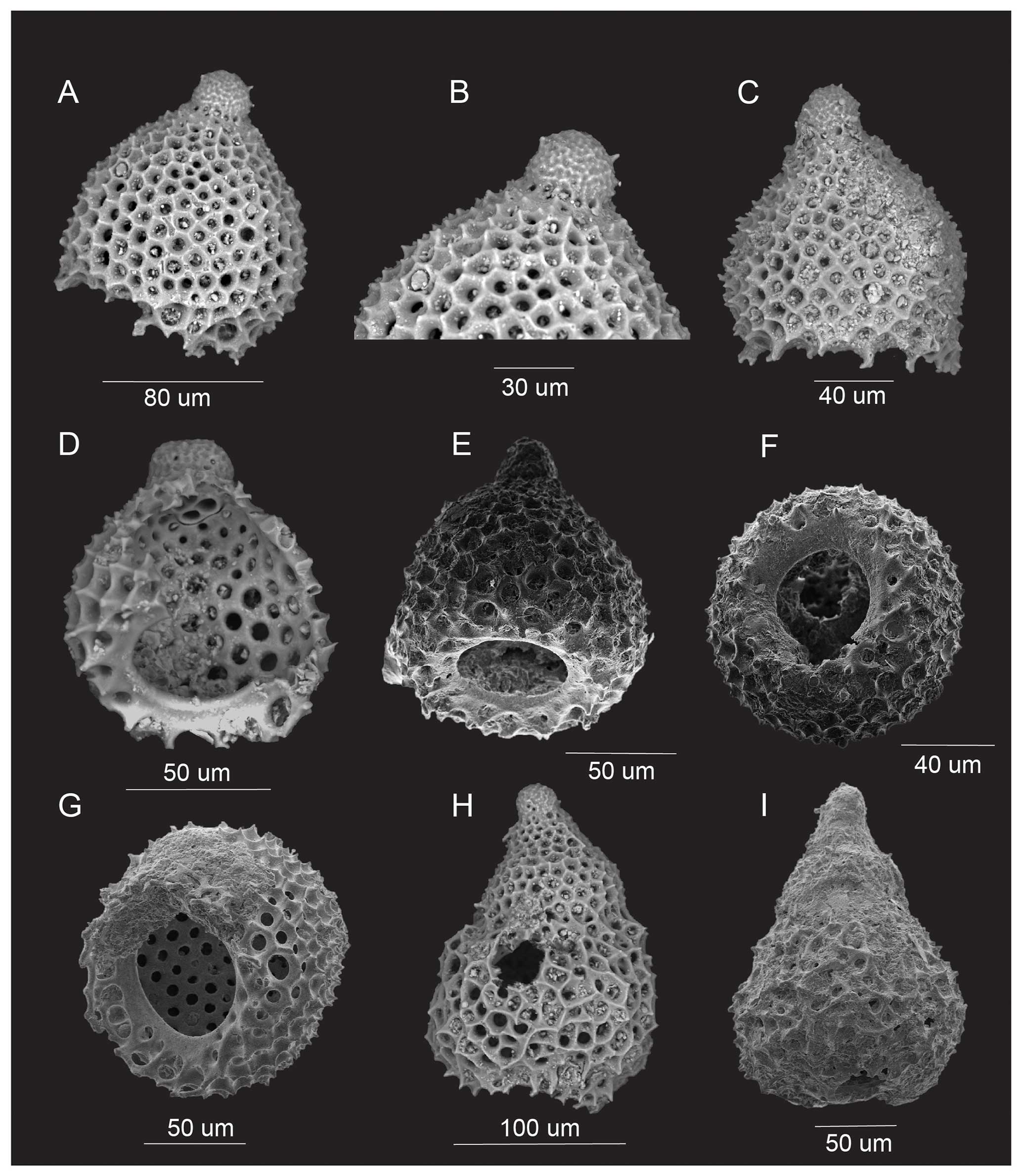
Plate 2(A–G) (sample C-628959) Kuppelella rusti n. sp. (A–B holotype number GSC-143277): (A) lateral view; (B) lateral view, zoom-in of cephalis; (C, E) lateral view; (D) lateral view showing internal structures and collar pores; (F–G) apertural view. (H–I) Lithocampe? rara n. sp. (H holotype number GSC-143278): sample C-628959, lateral view.
-
Superfamily Eucyrtidioidea Ehrenberg, 1846 sensu Suzuki in Suzuki et al., 2021
-
Family Eucyrtidiidae Ehrenberg, 1846 sensu Suzuki in Suzuki et al., 2021
-
Genus Lithocampe Ehrenberg, 1839
-
Lithocampe? rara n. sp. Diaz et Suzuki
(Plate 2, figs. H–I)
Derivation of name. From the Latin adjective “rara” (rare), after the uncommonly irregular arrangement of pores on the postabdominal segment.
Diagnosis. Four-segmented nassellarian with small, spherical cephalis and trapezoidal, perforate thorax. Post-abdominal segment large in absolute size. Pore frames raised with spine-like protuberances at vertices.
Holotype. Plate 2, fig. H, sample C-628959, deposited in the National Collection of Invertebrate and Plant Fossil Types in Ottawa, Canada, under repository number GSC-143278.
Material. Two specimens recovered from one sample (C-628959) in the uppermost part of the Smoking Hills Formation.
Range. Late Campanian.
Description. Test largely composed of four segments. Cephalis very small, spherical, without a horn, poreless or with few scattered pores. Collar stricture is well defined externally. Thorax is small in absolute size, trapezoidal and perforate. Thoracic pores are circular to subcircular, surrounded by pentagonal (?) frames. Lumbar stricture is poorly defined. Abdomen is slightly inflated, dome-shaped, perforate and small in absolute size. Stricture between abdomen and the postabdominal segment is well defined by abrupt change in size of postabdominal segment. Postabdominal segment is large in absolute size (more than half of the size of the whole test), perforate and subcylindrical tapering downwards into possible aperture. Pores on the post-abdominal segment are circular, of similar size and longitudinally arranged in some areas but irregular in others. Pores frames in the test, except for the cephalis, are raised with spine-like protuberances at vertices.
Dimensions (based on two specimens). Total height of test: 177 µm; width of cephalis: 22 µm; height of thorax: 19 µm; maximum width of thorax: 42 µm; diameter of the polygonal rimmed pore frame: 6–15 µm; number of pores in the half hemisphere of thorax: 7; number of pores in a vertical line of thorax: 5; width ratio of cephalis to thorax: 1.9; size ratio of pore to the maximum width of thorax: 0.24.
Remarks. This species is placed tentatively within the genus Lithocampe since the apertural end is not fully preserved in the few specimens recovered in this study. This species differs from other Lithocampe species in having a poorly defined lumbar stricture, partially irregular pore frame arrangement in the post-abdominal segment and a more constricted aperture.
The abundance and diversity of radiolarians in high northern latitudes are low compared to the mid and low latitudes (Empson-Morin, 1981), but they still allow for regional and sometimes global biostratigraphic correlations. Different species of Kuppelella Empson-Morin, 1981, for example, have been recovered from different parts of the world (e.g., Italy (Squinabol, 1903); the Pacific Ocean and coast of California (Empson-Morin, 1981; Campbell and Clark, 1944); western Siberia (Kozlova and Gorbovetz, 1966); India (Garg and Jain, 1978)) and could be used as markers for global biostratigraphic correlations. The species Canadasphaera inuita and Lithocampe? rara, on the other hand, seem to be restricted to the Arctic (as far as is known), and thus they may be more useful in regional studies. The Smoking Hills and Mason River formations have been dated as Santonian–Maastrichtian based on diatoms (Tapia and Harwood, 2002), silicoflagellates (McCartney et al., 2011) and dinoflagellates (McIntyre, 1974). The Campanian age of the Smoking Hills Formation is supported by the presence of exclusively Campanian radiolarian species such as Kreuzstella vierkantiga Empson-Morin, 1981, and Lithostrobus borealis Kozlova and Vishnevskaya, 2012 (Fig. 2). The upper Mason River Formation may be as young as Late Maastrichtian–Paleocene?, but the absence of siliceous microfossils and foraminifera in this interval prevents any definite age determination.
The absence or low quantity of radiolarians in Cretaceous strata from high northern latitudes could be related to poor living conditions during deposition or poor preservation. The few species recovered from Arctic regions typically exhibit poor preservation, challenging taxonomic descriptions and accurate classification. The preservation of the radiolarians recovered from the Smoking Hills and Mason River formations is moderate to good, depending on the stratigraphic position (Diaz et al., 2020). The species Kuppelella aff. cayeuxi (Squinabol, 1903), Phaseliforma concentrica (Lipman, 1952) and Dictyomitra multicostata Zittel, 1876, which were recovered almost exclusively from the Smoking Hills Formation (Fig. 2), are moderately preserved, and some external features are obscured by quartz recrystallization. Conversely, Canadasphaera inuita, Kuppelella rusti and Lithocampe? rara, recovered from the uppermost part of the Smoking Hills Formation and the lower and middle part of the Mason River Formation, exhibit good to exceptional preservation. The good preservation of radiolarians correlates well with the first appearances of diatoms, silicoflagellates and sponge spicules in the stratigraphic column (Tapia and Harwood, 2002; McCartney et al., 2011; Diaz et al., 2020). This direct relationship suggests that the preservation of radiolarians was enhanced by the presence of other siliceous microorganisms that may have caused oversaturation of silica in the seawater and/or at the water–sediment interface.
The mechanical rather than chemical methodology used in this study to recover the radiolarian shells and the dissection of those shells using razor blades and needles were key to describing and identifying radiolarian taxa. A thorough examination of the medullary shell of Canadasphaera inuita or the collar pores in Kuppelella rusti was only possible when some specimens were cut in half and then examined using an SEM. This technique is laborious and may destroy valuable specimens, but it proves helpful in refining the taxonomy of radiolarian taxa that have only been classified using their external morphology.
The taxonomic notes provided in this study serve as a foundation for future Cretaceous radiolarian research in Arctic regions. Generation of a formal Cretaceous radiolarian biostratigraphic zonation for the Arctic requires refinement in the taxonomy of these microfossils in areas where they have been reported before (e.g., the Schrader Bluff Formation in Alaska; the Niobrara, Lea Park, Pierre Shale, and Bearpaw formations in the interiors of Canada and the United States; and the Kanguk Formation in the Sverdrup Basin). There is also a need to correlate the few Upper Cretaceous radiolarian assemblages recovered in North America with the ones recovered in the Russian Arctic (e.g., Kozlova and Gorbovetz, 1966; Amon, 2000; Sarkisova, 2007; Vishnevskaya, 2011; Vishnevskaya and Kozlova, 2012) so that a radiolarian biostratigraphic zonation can be proposed in this part of the world. The absence of calcareous fossils in Cretaceous strata from high northern latitudes makes siliceous microfossils, including radiolarians, an essential tool to solve geoscientific problems such as ocean connectivity, marine plankton migration, ocean circulation and age determination.
All specimens are held in the National Collection of Invertebrate and Plant Fossil Types in Ottawa, Ontario, Canada, under repository numbers GSC-143273, GSC-143277 and GSC-143278.
JFD processed all the samples, analyzed, described and named some radiolarian taxa; imaged all the specimens; and edited the figures. NS described and named some radiolarian taxa. JMG and MB collected the samples, measured the stratigraphic section, and provided information on the geology of the study area and taxonomy. All authors wrote and edited the paper.
The contact author has declared that none of the authors has any competing interests.
Publisher’s note: Copernicus Publications remains neutral with regard to jurisdictional claims made in the text, published maps, institutional affiliations, or any other geographical representation in this paper. While Copernicus Publications makes every effort to include appropriate place names, the final responsibility lies with the authors.
We gratefully acknowledge the management of Rod Smith, the field party chief who organized and led the fieldwork in 2018. Thanks to Stephen Grasby for his assistance in the field and help in collecting the samples. Helicopter support was provided by Canadian Helicopters and the excellent piloting of Matt Fricker. We thank Per K. Pedersen (University of Calgary) for his support and guidance during this and related studies on Cretaceous radiolarians from the Canadian Arctic. Charles Henderson (University of Calgary) is also thanked for providing the tools necessary to complete the micropaleontological analysis. Finally, we would like to thank the journal reviewers, Liubov Bragina and Špela Goričan, and the editor Taniel Danelian for their constructive comments and suggestions to improve the manuscript.
This research has been supported by the Geological Survey of Canada through the GeoMapping for Energy and Mineral Program, the Faculty of Graduate Studies at the University of Calgary, and the Ministry of Science of Colombia (MINCIENCIAS).
This paper was edited by Taniel Danelian and reviewed by Liubov Bragina and Spela Gorican.
Adl, S. M., Bass, D., Lane, C. E., Lukeš, J., Schoch, C. L., Smirnov, A., Agatha, S., Berney, C., Brown, M. W., Burki, F., Cárdenas, P., ČepiČka, I., Chistyakova, L., del Campo, J., Dunthorn, M., Edvardsen, B., Eglit, Y., Guillou, L., Hampl, V., Heiss, A. A., Hoppenrath, M., James, T. Y., Karnkowska, A., Karpov, S., Kim, E., Kolisko, M., Kudryavtsev, A., Lahr, D. J. G., Lara, E., Le Gall, L., Lynn, D. H., Mann, D. G., Massana, R., Mitchell, E. A. D., Morrow, C., Park, J. S., Pawlowski, J. W., Powell, M. J., Richter, D. J., Rueckert, S., Shadwick, L., Shimano, S., Spiegel, F. W., Torruella, G., Youssef, N., Zlatogursky, V., and Zhang, Q.: Revisions to the Classification, Nomenclature, and Diversity of Eukaryotes, J. Eukaryot. Microbiol., 66, 4–119, https://doi.org/10.1111/jeu.12691, 2019.
Amon, E. O.: Upper Cretaceous radiolarians of the Urals, in: Materialy Po Stratigrafii i Paleontologii Urala (Materials on Stratigraphy and Paleontology of the Urals), Inst. Geol. Geokhim. Ural. Otd. Ross. Akad. Nauk. Vyp., Ekaterinburg, 5, 209 pp., 2000.
Basov, I. A. and Vishnevskaya, V. S.: Upper Mesozoic stratigraphy of the Pacific Ocean, Nauka, Moskow, 200 pp., 1991 (in Russian, summary in English).
Bergquist, H. R.: Micropaleontology of the Mesozoic rocks of northern Alaska, Geological Survey, Professional Paper 302-D, 1–227, 1966.
Bergstresser, T. J.: Radiolaria from the Upper Cretaceous Pierre Shale, Colorado, Kansas, Wyoming, J. Paleontol., 57, 877–882, 1983.
Campbell, A. S. and Clark, B. L.: Radiolaria from the Upper Cretaceous of middle California, Geol. Soc. Am. Spec. Paper, no. 57, 1–61, pls. 1–8, https://doi.org/10.1130/SPE57-p1, 1944.
Cavalier-Smith, T.: Principles of protein and lipid targeting in secondary symbiogenesis: euglenoid, dinoflagellate, and sporozoan plastid origins and the eukaryote family tree, J. Eukaryot. Microbiol., 46, 347–366, https://doi.org/10.1111/j.1550-7408.1999.tb04614.x, 1999.
Cavalier-Smith, T.: The phagotrophic origin of eukaryotes and phylogenetic classification of Protozoa, Int. J. Syst. Evol. Micr., 52, 297–354, https://doi.org/10.1099/00207713-52-2-297, 2002.
Cavalier-Smith, T.: Protist phylogeny and the high-level classification of Protozoa, Eur. J. Protistol., 39, 338–348, https://doi.org/10.1078/0932-4739-00002, 2003.
Cavalier-Smith, T., Chao, E. E., and Lewis, R.: Multigene phylogeny and cell evolution of chromist infrakingdom Rhizaria: contrasting cell organisation of sister phyla Cercozoa and Retaria, Protoplasma, 255, 1517–1574, https://doi.org/10.1007/s00709-018-1241-1, 2018.
De Wever, P., Dumitrica, P., Caulet, J. P., Nigrini, C., and Caridroit, M.: Radiolarians in the sedimentary record, CRC Press, Amsterdam, 1–533, https://doi.org/10.1201/9781482283181, 2001.
Diaz, J. F. and Velez, M. I.: Foraminiferal biostratigraphy of the middle Turonian-late Santonian interval in southern Saskatchewan, Canada, B. Can. Petrol. Geol., 65, 366–386, 2017.
Diaz, J. F. and Velez, M. I.: Late Cretaceous radiolarians from a bentonite-rich interval at the base of the Niobrara Formation, southwestern Saskatchewan, Canada: biostratigraphic and paleoenvironmental implications, Can. J. Earth Sci., 55, 321–329, https://doi.org/10.1139/cjes-2017-0115, 2018.
Diaz, J. F., Galloway, J. M., Bringué, M., Pedersen, P. K., and Grasby, S. E.: Siliceous microfossils and agglutinated foraminifera from the Upper Cretaceous Smoking Hills and Mason River formations in the Horton River area, Northwest Territories, Geological Survey of Canada, Open File 8716, 1–21, 2020.
Dixon, J.: A review of Cretaceous and Tertiary stratigraphy in the northern Yukon and adjacent Northwest Territories, Geological Survey of Canada, Paper 92-9, 79 pp., 1992.
Dixon, J.: Mesozoic-Cenozoic stratigraphy of the northern Interior Plains and plateaux, Northwest Territories, Geological Survey of Canada Bulletin, 536, 56 pp., https://doi.org/10.4095/210800, 1999.
Dixon, J. and McNeil, D. H.: Upper Cretaceous and Tertiary stratigraphy on the Tuktoyaktuk Peninsula, Northwest Territories, Geological Survey of Canada, Open File Report, 5703, 24 pp., 2008.
Ehrenberg, C. G.: Über die Bildung der Kreidefelsen und des Kreidemergels durch unsichtbare Organismen. Abhandlungen der Königlich Preussischen Akademie der Wissenschaften zu Berlin (1838), 59–147, https://www.biodiversitylibrary.org/page/29017435 (last access: 21 April 2024), 1839.
Ehrenberg, C. G.: Über 2 neue Lager von Gebirgsmassen aus Infusorien als Meeres-Absatz in Nord-Amerika und eine Vergleichung derselben mit den organischen Kreide-Gebilden in Europa und Afrika, Bericht über die zur Bekanntmachung geeigneten Verhandlungen der Königlich Preussischen Akademie der Wissenschaften zu Berlin (1844), 57–97, https://www.biodiversitylibrary.org/page/11052757 (last access: 26 April 2024), 1844.
Ehrenberg, C. G.: Über eine halibiolithische, von Herrn R. Schomburgk entdeckte, vorherrschend aus mikroskopischen Polycystinen gebildete, Gebirgsmasse von Barbados, Bericht der königlichen preussichen Akademie der Wissenschaften zu Berlin, 382–385, https://www.biodiversitylibrary.org/page/11056086 (last access: 25 April 2024), 1846.
Ehrenberg, C. G.: Über den Tiefgrund des stillen Oceans zwischen Californien und den Sandwich-Inseln aus bis 15600’ Tiefe nach Lieutenant Brooke, Monatsberichte der Königlich Preussischen Akademie der Wissenschaften zu Berlin (1860), 819–833, https://www.biodiversitylibrary.org/page/36276074 (last access: 24 April 2024), 1861.
Ehrenberg, C. G.: Fortsetzung der mikrogeologischen Studien als Gesammt-Uebersichtder mikroskopischen Paläontologie gleichartig analysirter Gebirgsarten der Erde, mit specieller Rücksicht auf den Polycystinen-Mergel von Barbados, Abhandlungen der Königlich Preussischen Akademie der Wissenschaften zu Berlin (1875), 1–225, https://www.biodiversitylibrary.org/page/30148696 (last access: 21 April 2024), 1876.
Empson-Morin, K. M.: Campanian Radiolaria from DSDP Site 313, Mid-Pacific Mountains, Micropaleontology, 27, 249–292, 1981.
Foreman H. P.: Upper Maestrichtian Radiolaria of California, Special Papers in Paleontological Association, 3, 1–82, 1968.
Garg, R. and Jain, K. P.: Polycystine radiolaria from phosphatic nodules of the Uttatur Formation, south India. (Part 1): Nassellaria, Biological Memoirs, 3, 131–175, 1978.
Grasby, S. E., Percival, J. B., Bilot, I., Ardakani, O. H., Smith, I. R., Galloway, J., Bringué, M., and McLoughlin-Coleman, T.: Extensive jarosite deposits formed through auto-combustion and weathering of pyritiferous mudstone, Smoking Hills (Ingniryuat), Northwest Territories, Canadian Arctic – A potential Mars analogue, Chem. Geol., 587, 120634, https://doi.org/10.1016/j.chemgeo.2021.120634, 2022.
Haeckel, E.: Entwurf eines Radiolarien-Systems auf Grund von Studien der Challenger-Radiolarien, Jenaische Zeitschrift für Naturwissenschaft 15, 418–472, https://www.biodiversitylibrary.org/page/8700599 (last access: 26 April 2024), 1882.
Haeckel, E.: Über die Ordnungen der Radiolarien, Sitzungsberichte der Jenaischen Gessellschaft für Medicin und Naturwissenschaft, Jena, für das Jahr 1883, 18–36, 1884.
Kennedy, A. E. and Coe, A. L.: Development of the freeze–thaw processing technique for disaggregation of indurated mudrocks and enhanced recovery of calcareous microfossils, J. Micropalaeontol., 33, 193–203, https://doi.org/10.1144/jmpaleo2013-020, 2014.
Kozlova, G. E. and Gorbovetz, A. N.: Radiolarians of the Upper Cretaceous and Upper Eocene deposits of the West Siberian Lowland, Proceedings of the All Union Petroleum Scientific Research Institute for Geological Survey (VNIGRI), 248, 1–159, 1966 (in Russian).
Kozur, H. and Mostler, H.: Systemmatical review of the up to now described Triassic radiolarians, in: Morphology, ecology and evolution of radiolarians. Material from the IV symposium of European radiolarists EURORAD IV, edited by: Petrushevskaya, M. G. and Stepanjants, S. D., Akademiya Nauk SSSR, Zoological Institute, Leningrad, USSR, 114–123, 1984 (in Russian).
Leidy J.: Notices of remains of extinct vertebrated animals discovered by Professor E. Emmons, P. Acad. Nat. Sci. Phila., 8, 255–256, 1856.
Levine, N. D., Corliss, J. O., Cox, F. E. G., Deroux, G., Grain, J., Honigberg, B. M., Leedale, G. F., Loeblich, A. R., Lom, I. J., Lynn, D., Merinfeld, E. G., Page, F. C., Poljansky, G., Sprague, V., Vavra, J., and Wallace, F. G.: A newly revised classification of the Protozoa, J. Protozool., 27, 37–58, https://doi.org/10.1111/j.1550-7408.1980.tb04228.x, 1980.
Lipman, R. K.: Materialy k monograficheskomu izucheniyu radiolyarii verkhnemelovykh otlozhenii Russkoi Platformy (Data on the monographic study of the radiolarian of the Upper Cretaceous deposits of the Russian Platform), Trudy Vsesoyuznogo Nauchno-Issledovatelskogo Geologicheskogo Instituta (VSEGEI), Leningrad, Paleontologiya i Stratigrafiya, 24–51, 1952 (in Russian).
McCartney, K., Witkowski, J., and Harwood, D. M.: Late Cretaceous silicoflagellate taxonomy and biostratigraphy of the Arctic Margin, Northwest Territories, Canada, Micropaleontology, 57, 61–86, https://doi.org/10.47894/mpal.57.1.03, 2011.
McIntyre, D. J.: Palynology of an Upper Cretaceous section, Horton River, District of Mackenzie, NWT. Geol. Survey Canada, Paper 74-14, 1–57, 1974.
McNeil, D. H.: New foraminifera from the Upper Cretaceous and Cenozoic of the Beaufort-Mackenzie Basin of Arctic Canada, Cushman Foundation for Foraminiferal Research, Special Publication 35, 95 pp., 1997.
Muehlbauer, R., Kelly, D. C., Bamburak, J. D., and Nicolas, M. P. B.: Late Cretaceous (Santonian–Campanian) marine microfossils of the Manitoba escarpment, southwestern Manitoba, in: Report of Activities 2014, Manitoba Mineral Resources, Manitoba Geological Survey, 181–186, 2014.
Müller, J.: Einige neue bei St. Tropez am Mittelmeer beobachtete Polycystinen und Acanthometren, Monatsberichte der Königlich Preussischen Akademie der Wissenschaften zu Berlin (1858), 154–155, https://www.biodiversitylibrary.org/page/11071854 (last access: 21 April 2024), 1859.
Myhr, D. W.: Markers within Cretaceous rocks as indicated by mechanical logs from boreholes in the Mackenzie Delta area, Northwest Territories, in: Report of Activities, Part B; Geological Survey of Canada, Paper 75-lB, 267–275, https://doi.org/10.4095/104317, 1975.
O'Dogherty, L., Caulet, J.-P., Dumitrica, P., and Suzuki, N.: Catalogue of Cenozoic radiolarian genera (Class Polycystinea), Geodiversitas, 43, 709–1185, https://doi.org/10.5252/geodiversitas2021v43a21, 2021.
Pessagno, E. A.: Radiolarian zonation and stratigraphy of the Upper Cretaceous portion of the Great Valley Sequence, California Coast Ranges, Micropaleontology, special Publication 2, 1–95, 1976.
Plauchut, B. P. and Jutard, G. G.: Cretaceous and Tertiary stratigraphy, Banks and Eglinton Islands and Anderson Plain (NWT), B. Can. Petrol. Geol., 24, 321–371, 1976.
Pugh, A. T., Schröder-Adams, C. J., Carter, E. S., Herrle, J. O., Galloway, J., Haggart, J. W., Andrews, J. L., and Hatsukano, K.: Cenomanian to Santonian radiolarian biostratigraphy, carbon isotope stratigraphy and paleoenvironments of the Sverdrup Basin, Ellef Ringnes Island, Nunavut, Canada, Palaeogeogr. Palaeocl., 413, 101–122, https://doi.org/10.1016/j.palaeo.2014.06.010, 2014.
Russell, D. A.: Cretaceous vertebrates from the Anderson River NWT, Can. J. Earth Sci., 4, 21–38, 1967.
Rust, D.: Radiolaria from the Pierre Formation of north-western Manitoba, in: Contributions to Canadian Micropaleontology, Geological and Natural History Survey of Canada, Vol. 4, 101–110, 1892.
Sandin, M. M., Biard, T., Romac, S., O'Dogherty, L., Suzuki, N., and Not, F.: A morpho-molecular perspective on the diversity and evolution of Spumellaria (Radiolaria), Protist, 172, 1–25, https://doi.org/10.1016/j.protis.2021.125806, 2021.
Sarkisova, E. V.: New radiolarian species from the Upper Cretaceous-Lower Paleogene of the eastern slope of the northern Ural Mountains, Paleontol. J., 41, 501–505, 2007.
Schultz, L. G., Tourtelot, H. A., Gill, J. R., and Boerngen, J. G.: Composition and properties of the Pierre Shale and equivalent rocks, northern Great Plains region (No. 1064-B), US Govt. Print. Off., B1–B114, 1980.
Smith, I. R., Bringué, M., Bryant, R., Evans, D. J. A., Galloway, J. M., and Grasby, S. E.: Multidisciplinary study of Cretaceous, Neogene, and Quaternary stratigraphy, and bocannes and their associated mineralogy and hydrochemistry, Smoking Hills, Northwest Territories: GEM-2 Western Arctic Project, report of activities 2018, Geological Survey of Canada, Open File 8482, 40 pp., 2018.
Squinabol, S.: Le Radiolarie dei noduli selciosi nella scaglia degli Euganei, Riv. Italiana Pal., 9, 105–150, pls. 8–10, 1903.
Suzuki, N., O'Dogherty, L., Caulet, J.-P., and Dumitrica, P.: A new integrated morpho-and molecular systematic classification of Cenozoic radiolarians (Class Polycystinea) – suprageneric taxonomy and logical nomenclatorial acts, Geodiversitas, 43, 405–573, https://doi.org/10.5252/geodiversitas2021v43a15, 2021.
Tapia, P. M. and Harwood, D. M.: Upper Cretaceous diatom biostratigraphy of the Arctic Archipelago and northern continental margin, Canada, Micropaleontology, 48, 303–342, https://doi.org/10.2113/48.4.303, 2002.
Tappan, H.: Cretaceous biostratigraphy of northern Alaska, AAPG Bull., 44, 273–297, 1960.
Tyrrell, J. B.: Report on northwestern Manitoba with portions of the adjacent districts of Assiniboia and Saskatchewan, Geological Survey of Canada, Annual Report, Vol. 5, pt. E, 1E–235E, https://doi.org/10.4095/216052, 1892.
Vishnevskaya, V. S.: Possibility of Jurassic-Paleogene volcanogenic-siliceous formations in the northwestern margin of the Pacific, in: Geological Outline of Kamchatka and Koryak Highland, Nauka, Moscow, 8–16, 1988 (in Russian).
Vishnevskaya, V. S.: New radiolaria of the family Prunobrachidae from the uppermost Cretaceous of the eastern Polar Urals, Paleontol. J., 45, 370–378, 2011.
Vishnevskaya, V. S. and Kozlova, G. E.: Volgian and Santonian – Campanian Radiolarian Events of the Russian Arctic and Pacific Rim, Acta Palaeontol. Pol., 57, 773–790, 2012.
Wall, J. H.: Diatoms and radiolarians from the Cretaceous system of Alberta, a preliminary report, in: The Cretaceous System in the Western Interior of North America, Geological Association of Canada, Special Paper 13, 391–410, 1975.
Yorath, C. J. and Cook, D. G.: Cretaceous and tertiary stratigraphy and paleogeography of the northern interior plains, district of Mackenzie, Geological Survey of Canada, Memoir 398, 76 pp., 1981.
Yorath, C. J., Balkwill, H. R., and Klassen, R. W.: Franklin Bay and Malloch Hill map-areas, District of Mackenzie, Geological Survey of Canada, Paper 74-36, 42 pp., 4 sheets, map 1403A, scale 1 : 250 000, 1975.
Young, H. R. and Moore, P. R.: Composition and depositional environment of the siliceous Odanah Member (Campanian) of the Pierre Shale in Manitoba, Geological Society of America, Special Papers 287, 175–196, 1994.
Zittel, K. A.: Uber einige fossile Radiolarien aus der norddeutschen Kreide, Zeitschrift der Deutschen Geologischen Gesellschaft, Stuttgart, 28, 75–86, 1876.
Zonova, T. D., Kasintsova, L. I., and Yazykova, E. A.: Atlas of the main groups of Cretaceous fauna from Sakhalin, Nedra, St-Petersburg, 1–327, 1993.