the Creative Commons Attribution 4.0 License.
the Creative Commons Attribution 4.0 License.
 | 03 Jun 2026
| 03 Jun 2026
First report of modern planktonic foraminifera in the southeastern Arabian Gulf, with observations on the morphology and ecology of Neogallitellia vivans (Cushman)
Sinatrya D. Prayudi
Thomas Steuber
Michael A. Kaminski
This study reports the first documented occurrence of planktonic foraminiferal assemblages from core and grab samples collected from the United Arab Emirates (UAE) sector of the Arabian Gulf, specifically from offshore Abu Dhabi. Although planktonic foraminifera were found at only 4 out of 20 surveyed sites, representing less than 5 % of the total foraminiferal assemblage, their presence is notable. This finding demonstrates that planktonic foraminifera can be found far from their typical habitat near the oceanic inlet to the semi-enclosed Arabian Gulf – the Strait of Hormuz. The recovered material includes at least five species (three true planktonic foraminifera, one with a planktonic adult stage, and one incertae sedis): Neogallitellia vivans, Globigerinoides tenellus, and Turborotalita clarkei, as well as the benthic–planktonic species Tretomphalus bulloides and a planktonic taxon of uncertain origin described under the name Bachmayerella sp. The species N. vivans dominates in the studied samples, comprising over 90 % of the planktonic assemblage; however, their small size means that most specimens are not retained on a 125 µm sieve. This work is part of a broader project to re-examine localities in the southeastern Arabian Gulf that were previously studied for benthic foraminifera, as well as to explore areas further offshore, which were previously uncharted for their foraminiferal content (particularly for the presence of planktonic foraminifera).
- Article
(7134 KB) - Full-text XML
-
Supplement
(11252 KB) - BibTeX
- EndNote




Planktonic foraminifera are a group of marine biocalcifiers with a planktonic mode of life and a cosmopolitan distribution across the world's oceans (Schiebel and Hemleben, 2017). Living planktonic foraminifera were first described during the HMS Challenger expedition when the ship passed through the Atlantic Ocean (Wyville-Thomson, 1878) and were later reported in almost every corner of the Earth. In total, 50 extant morphospecies of planktonic foraminifera currently inhabit the modern ocean (Brummer and Kučera, 2022), extending from tropical to polar regions, with five biogeographical provinces (Hemleben et al., 1989).
The occurrence of planktonic foraminifera in the modern ocean or in the sedimentary record provides valuable proxies that can be used to infer multiple environmental aspects. One application is understanding depth (or paleodepth) using the planktonic–benthic (PB) ratio (van der Zwaan et al., 1990) or even estimating paleoproductivity (Berger and Diester-Haass, 1988), such as in the case of the Arabian Sea during the last 135 kyr (Ivanova et al., 2003). Additionally, because certain species have preferred ecological traits, their occurrence can be used as an indicator of productivity or nutrient-rich or nutrient-poor conditions (Schiebel et al., 2001), which are normally associated with upwelling (Naidu, 1990), as well as for salinity and other geochemical proxies (see Katz et al., 2010 for details).
The Arabian Gulf (also known as the Persian Gulf or simply the Gulf) is an epicontinental marginal sea situated within an arid to semi-arid climatic zone (Purser and Seibold, 1973), originating from a tectonic depression formed during the Pliocene to Pleistocene (Kassler, 1973). Spanning approximately 1000 km in length and 300 km in width, the basin covers an area of roughly 226 000 km2 and exhibits a distinct bathymetric asymmetry: the average bathymetrical depth of 35 m that deepens to nearly 100 m along the northern Iranian margin while shallowing gently towards the southern and western region (Saudi Arabia, the United Arab Emirates (UAE), Qatar, and Bahrain). Oceanographically, the Gulf is characterized by limited seawater exchange through the Strait of Hormuz as freshwater input is negligible and restricted primarily to the Shatt Al-Arab estuary in the northwest and minor Iranian tributaries (Sheppard et al., 1992). Incoming surface water from the Indian Ocean (±37 psu) follows an anti-clockwise circulation pattern, starting along the northern coast and moving westward before high evaporation rates drive a density increase, causing the water to sink towards the south and exit as a deep hypersaline flow (Sheppard et al., 1992, 2010). Consequently, while the basin is generally hypersaline (>40 psu), restricted areas with poor circulation can exceed 70 psu (Prayudi et al., 2024a; Gull et al., 2025), creating intense physicochemical conditions that, alongside anthropogenic pressures, act as controlling factors for the region's marine biota (i.e. temperature during summer; see Prayudi et al., 2024b).
Studies on the occurrence of planktonic foraminifera in the Arabian Gulf can be traced back to the 1970s. Initially, Hughes Clarke and Keij (1973) mentioned the presence of planktonic foraminifera in the deepest sector with non-restricted conditions of the basin, closer to the Strait of Hormuz and diminishing westward. Lutze (1974) provided an initial report on benthic foraminiferal assemblages (in addition to the presence of planktonic foraminifera) using sediment collected by a grab sampler. In that work, the author quantified the percentages of planktonic foraminifera from the deeper, Iranian side of the Arabian Gulf and observed values above 50 % in terms of the total foraminiferal assemblage. Lutze also noted the occurrence of the species “Guembelitria” vivans Cushman. Previous reports of this species suggest that the species is mostly found in the Indo-Pacific region (Fig. 1A–C), although some studies previously mentioned that, by looking at the smaller fraction (<125 µm), this species (and other minute-sized species) can contribute as the majority of the planktonic foraminiferal contents (Kroon and Nederbragt, 1990; Darling et al., 2009). We can infer the possibility that it is cosmopolitan in distribution. A subsequent report by Cherif et al. (1997) also found samples with planktonic foraminifera near the Strait of Hormuz, which serves as the only marine inlet. However, Cherif et al. (1997) did not find any specimens of N. vivans, and their planktonic foraminiferal relative abundances are not as high as at some of Lutze's sites (reaching up to 50 %). After the Cherif et al. (1997) study, no further reports on the occurrence of planktonic foraminifera within the Arabian Gulf have been published.
In this study, we report a new finding of planktonic foraminifera in the southeastern sector of the Arabian Gulf. The purpose of this report, which is part of a larger effort to analyse newly collected materials from this region (Steuber et al., 2026), is to document the distribution of planktonic foraminifera and to shed light on the ecological role of these biocalcifiers in the region. Specifically, this work will help establish a baseline to study the extended distribution of modern planktonic foraminifera in this restricted and environmentally stressed environment. We also examine the ultrastructural features of N. vivans, thereby adding new observations to our present knowledge of this taxon.
A total of 20 sediment samples were collected from two legs of an ocean expedition of the R/V Jaywun, conducted by means of collaboration between the Environmental Agency Abu Dhabi and Khalifa University of Science and Technology, Abu Dhabi (Fig. 1D–E). In addition to the sediment sampling, we measured the physical properties of the seawater during the expedition by utilizing Conductivity, Temperature, Depth (CTD) (Sea Bird™ SBE911plus and SBE19plus) to measure the temperature and salinity along the transect that we visited during the cruise. The first sampling event was performed in September 2024 (Steuber et al., 2026), collecting sediment cores at six stations using a Maxi Core device (OSIL Bowers & Connelly Multiple Maxi Corer, 110 mm diameter). The second sampling event was conducted in March 2025, retrieving 14 sediment samples using a Van Veen grab sampler (2000 cm2). No staining procedure was performed during the first expedition since the main purpose of the first (and also the second) expedition was to survey the sediment composition and to collect oceanographic information (Steuber et al., 2026). Samples from the second expedition were stained using rose Bengal mixture for a duration of 2 weeks, following the standard procedure by FOBIMO (Schönfeld et al., 2012), as the initial intention was to check for living benthic foraminifera in the retrieved samples (part of supplementary work dealing with the benthic foraminifera, Prayudi et al., 2026).
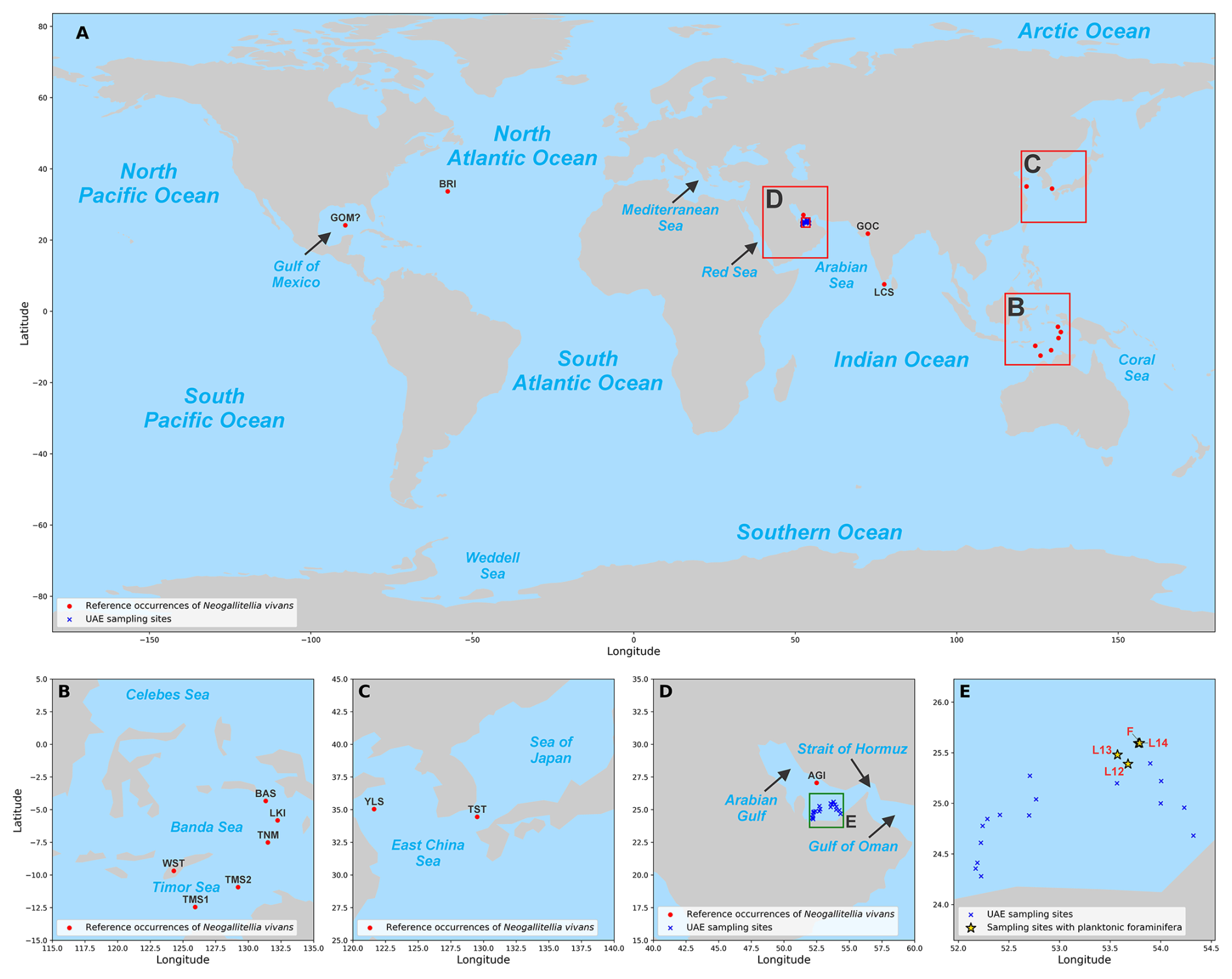
Figure 1Distribution maps showing the global occurrence of N. vivans (A–D) and specific sampling sites in this study (E). The sites in (E) correspond to locations where other planktonic foraminiferal species are also present (4 out of 20 sites). Abbreviations used in the maps, as well as in the source of the reference points, are as follows: GOM – Gulf of Mexico (Loeblich and Tappan, 1986); BRI – Bermuda Rise (Pawlowski, 1991); GOC – Gulf of Cambay (Ghosh et al., 2008, 2009; Ghosh, 2012); LCS – Laccadive Sea (Kroon and Nederbragt, 1990); WST – West Timor (De Smet et al., 1990); TMS – Timor Sea (Loeblich and Tappan, 1986); TNM – Tanimbar Island (Charlton et al., 1991); LKI – Little Ki (Kai) Island (Cushman, 1934); BAS – Banda Sea (Kroon and Nederbragt, 1990); YLS – Yellow Sea (Zheng and Fu, 2008); TST – Tsushima Strait (Ujiié et al., 2008; Kimoto et al., 2009); AGI – Arabian Gulf-Iran (Lutze, 1974).
Upon the preparation of the samples, around 30 g of sediments was washed with flowing water over a 63 µm sieve to remove mud and clay particles, as well as excessive stain. The wet samples were then dried in an incubator for 24 h at a temperature of 60 °C. To reduce the picking time while avoiding sampling bias, we split the dried samples using a Micropress Europe microsplitter until the subsample contained ca. 300 specimens (both benthic and planktonic), and then we further dry sieved the residues over a 125 µm sieve. The foraminifera were observed using Olympus SZX7 with Olympus KL 300 LED and then were picked using a 3/0 brush and transferred to cardboard microslides. Samples containing the highest relative abundance of planktonic foraminifera (sample L14) were picked in duplicate in order to ensure recovery of meaningful numbers of specimens and to verify their abundances (samples L14-1 and L14-2).
For documenting the detailed species identification and morphological features, we performed scanning electron microscopy (SEM) observations in two stages. In the first stages, for the general observation and taxonomical determination, we used a JEOL NeoScope JCM-7000, with specimen placement using carbon tape and coated with gold. In the second stage, for imaging the detailed morphological features such as pore measurements and ultrastructural observations, we used a Zeiss Gemini SEM 450 with the same procedure for sample preparation (placement and coating). In both observations, we used 15 kV electron beam penetration in order to observe the morphology of the foraminiferal tests, resulting in higher resolution in the final images.
As this study targets the planktonic foraminifera, we used several reference monographs, reports, and publications to determine our recovered species. We utilized Schiebel and Hemleben (2017) and Brummer and Kučera (2022) as our guide for identifying modern planktonic foraminifera and Loeblich and Tappan (1987) for general foraminiferal taxonomical descriptions, as well as the subsequent literature to chart the occurrence of N. vivans elsewhere in the world ocean (see Fig. 1A–D and the provided references in the figure caption).
To contextualize the global distribution of N. vivans and to map the spatial abundance of planktonic foraminifera within the Arabian Gulf, we generated a series of maps using Python (Spyder 6 environment). For the global and regional occurrence maps, geographical coordinates of past reports and our new sampling sites were compiled and projected using the pandas and geopandas libraries. The underlying land and ocean boundaries were sourced from the Natural Earth 1:110m physical vectors dataset.
For the spatial abundance maps within the Arabian Gulf, we compared our findings with 121 historical sampling coordinates extracted from the Amao et al. (2025) repository, which digitized the Lutze (1974) dataset. To visualize the spatial gradients of both the total planktonic foraminifera and N. vivans percentages, we applied a linear interpolation over a high-resolution regular grid using the griddata function from the scipy.interpolate library. The resulting interpolated grid was then geographically masked using an Arabian Gulf shapefile to restrict the visualized contours strictly to the marine basin and was finally plotted using matplotlib.pyplot.
3.1 Overview of the physical ocean parameters from the sampling sites
Seawater temperature values, as measured with Sea Bird™ SBE911plus and SBE19plus CTDs, were as high as 35.6 °C in August 2024 in the southwestern part of the study area and were uniformly above 35 °C throughout the water column for 200 km offshore, down to 35 m of water depth. Further offshore, at the sites from which planktonic foraminifers are reported here (Fig. 1E), and approaching the main basin of the Gulf, sea surface temperature was similar but dropped to 26 °C at 60 m depth.
In March 2025, the coastal and sea surface water had already started to warm from the winter minimum, and the lowest observed temperature was 21 °C at a depth of >20 m in the restricted basin to the south of the Great Pearl Bank (see Steuber et al., 2026, for further details).
Salinity of 44 to 45 psu throughout the water column was observed with CTD surveys in the basin south of the Great Pearl Bank in August 2024 and March 2025. Further to the east of the study area, salinity reached 42 psu in summer 2024 and 43 psu during March 2025 in coastal bottom waters at 20 m depth. In both surveys, salinity of surface waters dropped to 39 psu further offshore, i.e. at the sampling sites of the planktonic foraminifera (Fig. 1E), showing the influx of surface water with a lower salinity from the central basin of the Gulf, approaching the values of the surface inflow from the Arabian Sea of 37 psu (Reynolds, 1993; Steuber et al., 2026).
3.2 Overview of the observed species
In the sediment samples provided from offshore Abu Dhabi, there are four localities, one from the first sampling campaign (sample F) and three sites from the second campaign (samples L12, L13, and L14), that contain planktonic foraminifera (Fig. 1E). In the duplicate samples that we picked, we found similar occurrences of planktonic foraminifera in both sample splits. The percentages of planktonic foraminifera for each sites range from 0.12 % to 5.22 % from the total picked foraminiferal specimens (relative to the benthic foraminifera or total assemblages). The collected materials were based on seafloor sediments; hence, the planktonic foraminifera specimens were already dead. The stained samples collected during the second campaign confirmed this assumption.
We found, in total, three species of true planktonic foraminifera (Plate 1), namely Globigerinoides tenellus Parker, 1958, Turborotalita cf. clarkei (Rögl & Bolli, 1973), and Neogallitellia vivans (Cushman, 1934), as well as one species, Tretomphalus bulloides (d'Orbigny, 1839), that, according to Banner et al. (1985), can be considered to be transitional between benthic and planktonic owing to the presence of a planktonic phase during its life cycle. Among all planktonic species, N. vivans is the most abundant species. The other records are just single occurrences, with a maximum of three counts per species in the studied samples, and not all samples contain similar occurrences of planktonic foraminifer (Tables 1 and 2).
Additionally, we found one possible planktonic shell of uncertain origin (incertae sedis), which only occurs in one sample (L14-2). This taxon is characterized by its spherical morphology, with multiple pores that are scattered over the shell or outer layer (referring to Versteegh et al., 2009). It resembles the spherical planktonic foraminifera Orbulina universa but is morphologically distinctive in terms of the absence of apertures and its smaller size. The morphologically closest form is Bachmayerella sp., a microfossil of uncertain origin initially described by Rögl and Franz (1979) from the Miocene of the Paratethys.
Table 1Sampling sites and associated planktonic foraminiferal abundances recorded in this study. Note that “PF” denotes planktonic foraminifera.
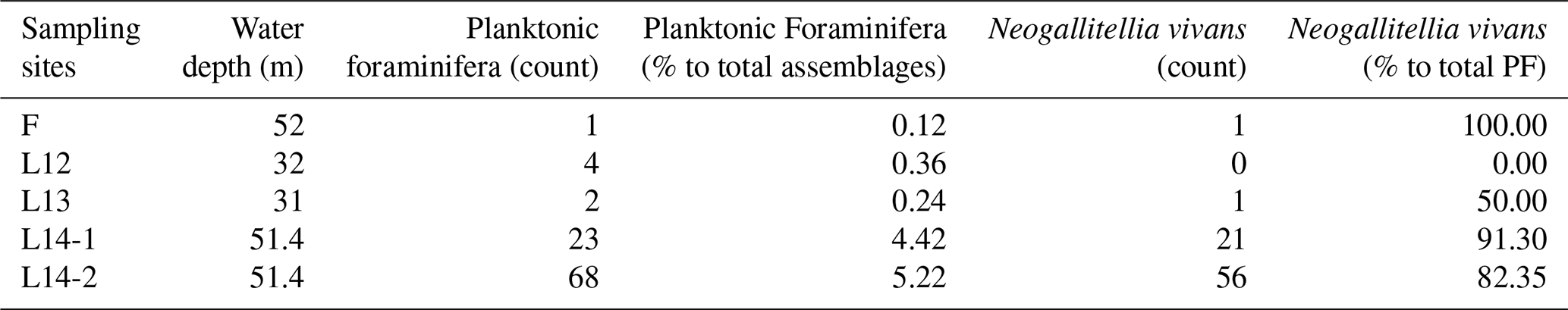
Table 2List of planktonic foraminiferal species and associated species and their occurrence at each sampling site. Note that “PF” denotes planktonic foraminifera.
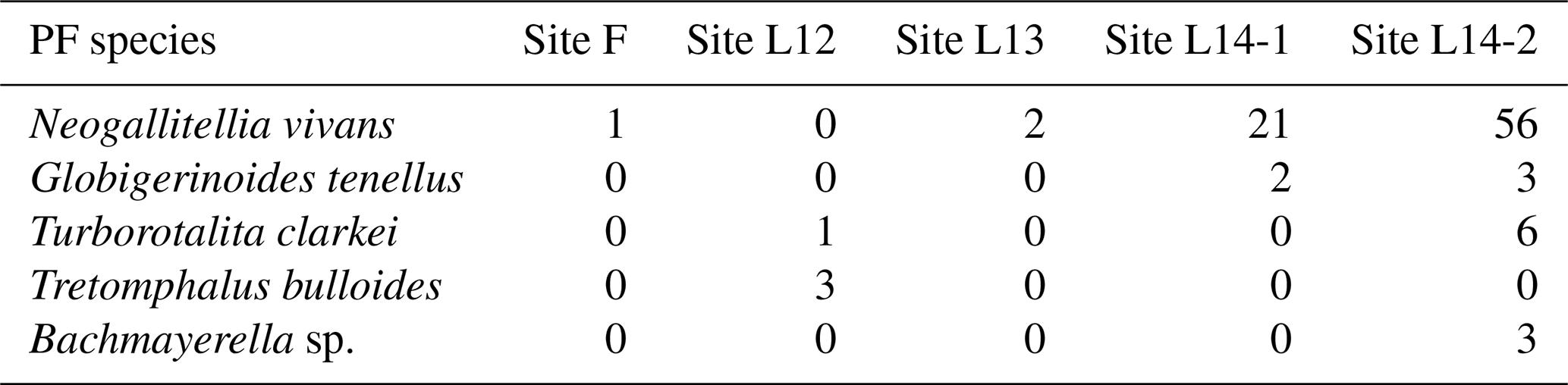
Most of the specimens were found in samples collected from the deeper stations on the UAE shelf (sample L14). Even though this species is well known for its small size (less than 120 µm) and possibly passes through the 125 µm sieve, some of the specimens that are retained on the sieve are quite large (Table 1, Fig. 2).
3.3 Systematic taxonomy – a redescription of N. vivans (Cushman)
-
Order ROTALIIDA Lankester, 1885
-
Suborder GLOBIGERININA Delage and Hérouard, 1896
-
Superfamily GUEMBELITRIOIDEA Montanaro Gallitelli, 1957
-
Family GUEMBELITRIIDAE Montanaro Gallitelli, 1957
-
Genus Neogallitellia Özdikmen, 2009
Type species. Guembelitria vivans Cushman, 1934
Remarks. Neogallitellia Özdikmen, 2009 is a replacement name for Gallitellia Loeblich and Tappan, 1986, which was a junior homonym of the anthozoan Gallitellia Cuif, 1977.
-
Neogallitellia vivans (Cushman, 1934)
Plate 1, figs. 1–3; Plate 2, figs. 1–3.
-
1934 Gümbelitria(?) vivans Cushman, p. 105, pl. 13, figs. 9–10.
-
1974 Gümbelitria vivans Cushman, 1934. Lutze, p. 45, pl. 11, fig. 177.
-
1986 Gallitellia vivans (Cushman, 1934). Loeblich and Tappan, p. 249, figs. 9–12.
-
1990 Gallitellia vivans (Cushman, 1934). Kroon and Nederbragt, p. 31, pl. 1, figs 1–10.
-
1991 Gallitellia vivans (Cushman, 1934). Pawlowski, 1991, pl. 1, fig. 4.
-
2009 Gallitellia vivans (Cushman, 1934). Kimoto et al., p. 74, figs. 3, 4.
-
2009 Neogallitellia vivans (Cushman, 1934). Özdikmen, p. 241.
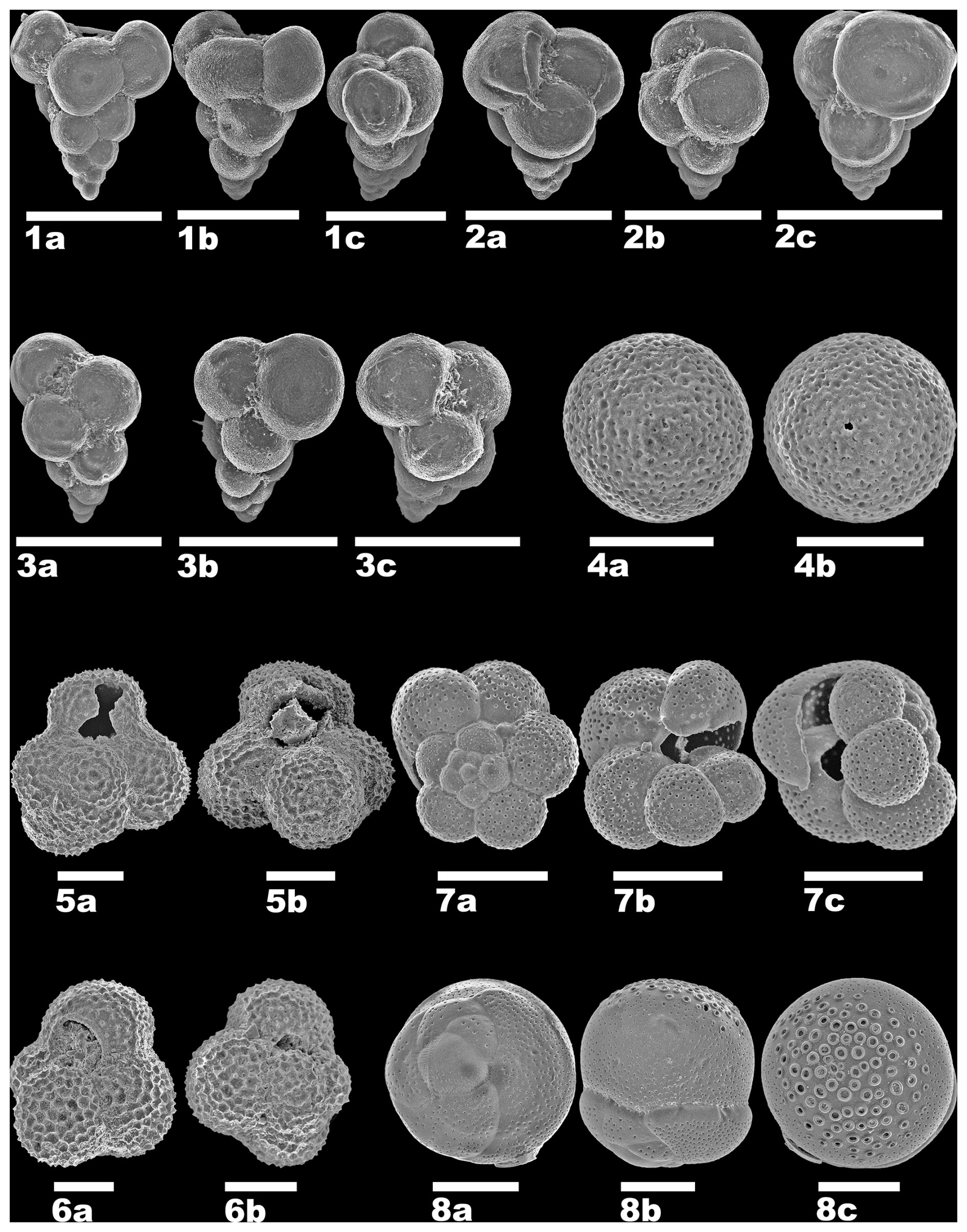
Plate 1Planktonic foraminifera and associated species in the southeastern Arabian Gulf. (1–3) Neogallitellia vivans (Cushman, 1934), site L14-2. (4) Bachmayerella sp., site L14-2. (5) Globigerinoides tenellus Parker, 1958, site L14-2. (6) Globigerinoides tenellus Parker, 1958, site L14-1. (7) Turborotalita cf. clarkei (Rögl & Bolli, 1973), site L12. (8) Tretomphalus bulloides (d'Orbigny, 1839), site L12. All scale bars are 100 µm.
Material. 79 specimens.
Description. Test tiny (averaging 130 µm based on the picked specimens retained on a 125 µm sieve), elongate, chambers globular and rapidly enlarging as added, arrangement triserial throughout, with chamber proliferation occasionally observed in the final stage, sutures deeply depressed, margins lobulate; wall calcareous, hyaline, consisting of a single continuous layer, microperforate, thin and translucent, surface smooth to grainy in appearance, with fine pores (<0.5 µm) randomly and sparsely scattered over the surface, often occurring in doublets or triplets, not elevated on pore mounds or in pore pits; aperture a simple rounded and umbilical arch at the base of the final chamber, without a lip.
Remarks. Cushman (1934) described the species from Challenger Station 192A off Little Ki Island, Indonesia, at 129 fathoms. Cushman's illustrated paratype shows strong and irregular chamber proliferation in the final growth stage. Specimens from the Arabian Gulf may become terminally quadriserial (Plate 1, fig. 1a–c), but we have not observed such irregular chamber proliferation in our material.
Our investigation of un-etched broken specimens (Plate 2, figs. 1–3, sample S1a–S3c) reveals that the test wall is very thin and consists of a single, contiguous calcitic layer lacking a visible primary organic sheet, with randomly (not regularly) spaced micropores, less than half a micron in diameter. This feature was previously displayed by Kroon and Nederbragt (1990) in Plate 1, fig. 9, but was not mentioned explicitly, and it was later observed by Kimoto et al. (2009, Fig. 3) from light microscope observation. Pores are sparse or absent in the proximity of the sutures (Plate 2, fig. 1b) and often occur as closely spaced pairs (doublets) or triplets in a triangular arrangement (Plate 2, figs. 1b, 2b, and 3b). This characteristic was also previously displayed in the SEM images of Loeblich and Tappan (1986, figs. 9–12), Kroon and Nederbragt (1990, plate 1, figs. 5 and 8), Pawlowski (1991, plate 1, fig. 4), Ujiié et al. (2008, fig. 2), Kimoto et al. (2009, fig. 4), and Schiebel and Hemleben, (2017, plate 2.32) but was not mentioned or emphasized in greater detail. The surface of the test is smooth, without pore mounds or pits.
According to Loeblich and Tappan (1986), the genus differs from Guembelitria Cushman and Chiloguembelitria Hofker (= Jenkinsina Haynes) in terms of its very thin, hyaline, smooth, and sparsely perforate wall; the occasional chamber proliferation; and the simple open aperture that has no bordering lip. It also differs from Guembelitria in terms of the wall lacking pore mounds. The mid-Cretaceous Guembelitriella also has chamber proliferation but differs in its finely pustulose surface, with pores between the pustules, and in the low arched aperture with a bordering lip.

Plate 2Detailed morphological and ultrastructural observations of N. vivans. Three broken specimens (1–3) are displayed to illustrate various morphological features: (a) complete specimens, with partial breakage to reveal the wall structure; (b) surface pore patterns and general morphological characteristics; and (c) side-view observations of pores and wall layering in the middle and final chambers.
Distribution. The specimens recovered from offshore the southeastern Arabian Gulf were found in sampled sediments at a water depth of 35–40 m, where it was the most abundant planktonic foraminiferal species. Neogallitellia vivans is regarded to be a shallow-dwelling epipelagic species – Kroon and Nederbragt (1990) collected living specimens from a water depth of 0–5 m off the southern tip of India, and Kimoto et al. (2009) recovered living specimens from plankton tows between 1 and 30 m water depth in the Tsushima Strait, at the entrance to the Japan Sea (see Fig. 1). Pawlowski (1991) reported a sediment sample with common N. vivans dated to 11 900 ka in core EN190-GGC1 from the Bermuda Rise. Kimoto et al. (2009) regarded the species to be adapted to warm, nutrient-rich coastal waters, noting that it was absent in the colder waters of the Japan Sea. The common occurrence of Neogallitellia offshore Abu Dhabi suggests that the species tolerates a wide range of salinities, including the hypersaline conditions of the southeastern Arabian Gulf.
3.4 Overview of general distribution of the targeted taxa
From the extracted and later interpolated datasets, the general distribution pattern from both aspects is relatively clear (Fig. 2). For the planktonic foraminifera, the highest percentages (up to 50 %) are found in the eastern part of the Arabian Gulf, and their relative abundance decreases westward (close or moving to 0). On the other hand, the percentage of N. vivans tends to be higher in the inner part of the Gulf, particularly in the central part (including in our sites, where values exceed 50 %), and is reduced towards the northeast and northwest.
4.1 Significance of planktonic foraminiferal occurrence in the southeastern Arabian Gulf
The presence of planktonic foraminifera in the inner Arabian Gulf is not well documented and established, with only sporadic reports over the last 50 years, with almost 3 decades of no reports (1998 to 2025). Hughes Clarke and Keij (1973) reported occurrences as far west as eastern Qatar, though most were concentrated near the Indian Ocean inlet. Lutze (1974) and Cherif et al. (1997) identified several species near the Strait of Hormuz. Cherif et al. (1997) classified the Abu Dhabi region as “Group 4 Southern (Arabian) Shallow Shelf”, noting that planktonic foraminifera were either absent or present in unspecified, negligible quantities. However, no studies have previously documented the occurrence of planktonic foraminifera along the southeastern coast, specifically from the United Arab Emirates and particularly from offshore Abu Dhabi. Therefore, this study presents the first report of their occurrence in this region, expanding current knowledge regarding their distribution within the modern-day Arabian Gulf.
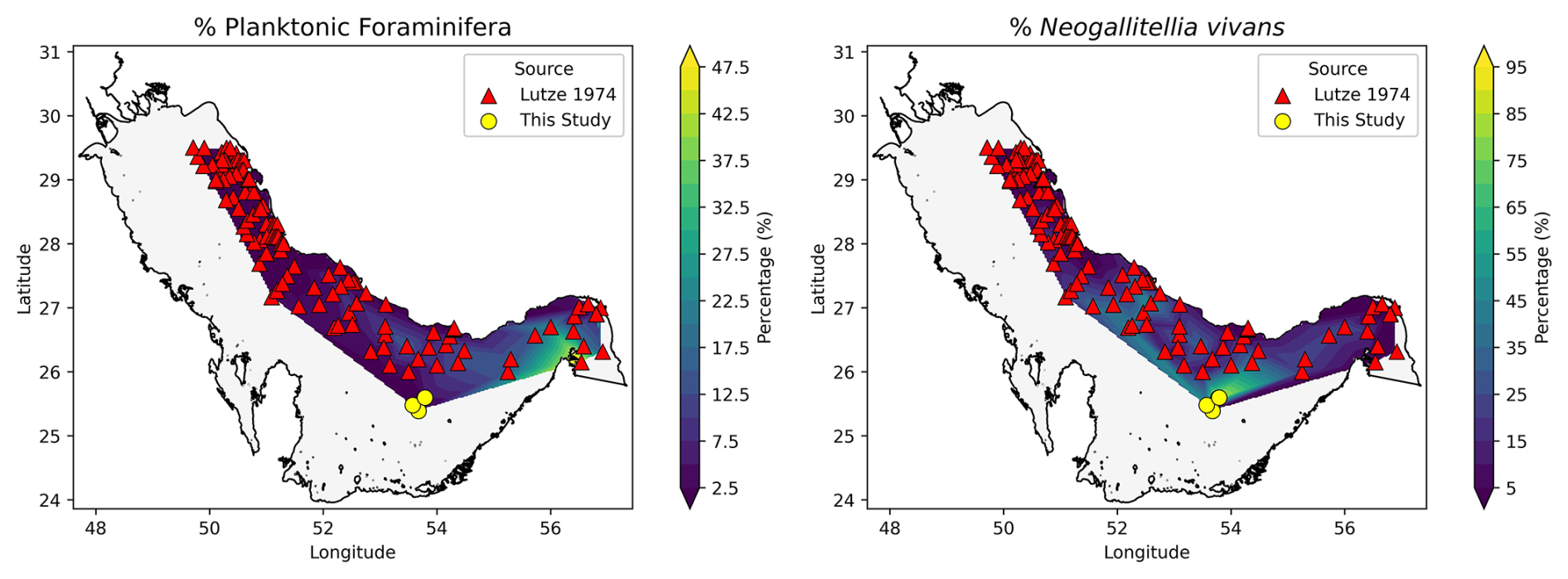
Figure 2Distribution maps of planktonic foraminifera (left) and the targeted species (right). Interpolated data originate from Lutze (1974) and are further augmented by our new samples.
From the distribution maps provided from the interpolated data, we infer that planktonic foraminifera in the Arabian Gulf originate from the Indian Ocean. As shown by Hughes Clarke and Keij (1973) and in the combined datasets of Lutze (1974) and our sites, the general pattern of planktonic foraminifera shows higher numbers closer to the inlet on the eastern side of the Arabian Gulf (Strait of Hormuz), with abundances decreasing westward (Fig. 2). This finding corroborates the additional report by Cherif et al. (1997) and aligns with general ideas about the presence of planktonic foraminifera in the Gulf of Oman and the northern Arabian Sea (Brock et al., 1992; Schiebel et al., 2004).
There are several mechanisms that might explain the presence of planktonic foraminifera in a semi-isolated, shallow marine basin like the Arabian Gulf, even as dead assemblages, despite their true nature as a plankton organism. One plausible explanation is that surface currents from the Indian Ocean might transport planktonic organisms, including foraminifera and calcareous nannoplankton, into the basin (Subba Rao and Al-Yamani, 1998; Vasou et al., 2020; Steuber et al., 2026). Alternatively, another explanation could be invasion from outside the basin through the introduction of ballast water, a known vector for invasive marine species in this region due to heavy commercial shipping (Al-Yamani et al., 2015). A third possibility is that planktonic foraminifera are simply part of a viable plankton community that had established itself in the deepest sectors of the Gulf at some stage during the Holocene, during the postglacial reflooding of the marine basin (Lambeck, 1996; Alsharhan and Kendall, 2003; Sheppard et al., 2010). Particularly in the case of the enigmatic genus Bachmayerella, this taxon was quite common in a semi-isolated setting within the Paratethys (Spezzaferri and Rögl, 2004), which was, in many ways, similar to the modern-day Arabian Gulf in terms of its paleoenvironment and paleoecological parameters (temperature and salinity context; see Schenk et al., 2018; Kranner et al., 2021; Peryt et al., 2024).
However, there is currently insufficient evidence to validate any of these hypotheses (apart from the last one, partially, which will be discussed below). Data on the spatio-temporal variation and historical record of foraminiferal assemblages in the Gulf's deeper sections are either unavailable or unexplored, with limited accessibility due to the proximity of offshore oil platforms (Hamza and Munawar, 2009). Furthermore, existing plankton studies have focused on other meiofaunal groups, such as copepods (Liu et al., 2022), or were conducted in the adjacent Arabian Sea rather than in the Arabian Gulf itself (e.g. Peeters et al., 1999; Schulz et al., 2002).
4.2 Neogallitellia vivans occurrence and redescription: overlooked aspects and their significance
The presence and dominance of N. vivans in our samples support the hypothesis that this species functions as a stress-tolerant “disaster taxon” within the Arabian Gulf since the basin itself mostly cited as a naturally and anthropogenically stressed environment (Sheppard et al., 1992, 2010). In the fossil record, minute, triserial planktonic foraminifera have thrived during extreme environmental crises, such as the K–Pg boundary event (Keller, 2003; Keller and Pardo, 2004; Pardo and Keller, 2008). Additionally, this is also the case for the extant biserial form Streptochilus, which displays blooms even in oceanic settings.
This hypothesis is consistent with the geological history of the Arabian Gulf, a relatively young marine basin (Sheppard et al., 2010) formed by reflooding during the early-Holocene transgression (see Lambeck, 1996). Although modern hypersaline conditions differ from the oceanic conditions after the Cretaceous–Paleogene extinction event, the persistence of adaptive traits in N. vivans is highly plausible. The species appears to function as an opportunistic niche occupier in environments where typical planktonic foraminifera cannot survive (Darling et al., 2009; Schiebel and Hemleben, 2017). This interpretation is supported by Lutze (1974), who observed that, while standard planktonic assemblages comprise up to 50 % of the seafloor fauna near the Strait of Hormuz, N. vivans dominates the deepest region of the northern Arabian Gulf, which was the first to be submerged during the Holocene reflooding (Lambeck, 1996; Alsharhan and Kendall, 2003). Consequently, as the only extant triserial form in the modern ocean, N. vivans, appears to be uniquely adapted to flourish in eutrophic and hypersaline coastal environments that do not support other planktonic foraminiferal species. Future work needs to check for the presence of living specimens in plankton samples collected from the region.
With regard to the significance of this taxon's morphology, our un-etched SEM cross-sections consistently reveal a thin, single-layered wall structure. This structural simplicity aligns with the previous observations of Kimoto et al. (2009), who suggested a possible monolamellar wall structure for this species. In addition, we performed classical yet detailed microstructural observations on broken specimens, similarly to what Hemleben (1969) carried out for his observations of molamellar structure in Hastigerina pelagica. While microperforate taxa are classically defined as bilamellar (Schiebel and Hemleben, 2017), our specimens present a structural simplicity that resembles a monolamellar wall structure, aligning with the previous light-microscope observations of Kimoto et al. (2009). We report this single-layered appearance strictly as a morphological observation of our regional specimens rather than as an attempt to definitively revise the higher-level taxonomy of the clade. In the meantime, we point out that future ultrastructural studies utilizing chemical etching protocols would be required to definitively confirm the presence or absence of a primary organic sheet; these distinctive wall features and pore patterns likely reflect its evolutionary and ecological background.
Regarding the pore pattern, foraminiferal porosity is primarily functional for gas exchange, with pore size and density often serving as proxies for ambient oxygen levels (Glock et al., 2012). This is consistent with the ecology of N. vivans, which is frequently associated with upwelling regions and oxygen-minimum zones (OMZs), such as those in the Arabian Sea (Kroon and Nederbragt, 1990; Ghosh et al., 2008). Phylogenetically, Ujiié et al. (2008) indicated that N. vivans diverged from its benthic ancestor Stainforthia fusiformis during the late early Miocene (∼ 18.2 Ma). This species (S. fusiformis) is characterized by minute pores (Hofker, 1956; Williamson, 1858), and its initial chambers bear a strong resemblance to those of N. vivans. Stainforthia is well-known for its tolerance of dysoxic environments (Alve, 2003). We suggest that these retained traits are linked to the species' transition from a benthic to a planktonic lifestyle, similarly to the extant biserial forms (see Darling et al., 2009), though this aspect is beyond the scope of the current study, and further investigations are needed to observe its living behaviour, such as laboratory culture and/or micro-mesocosm experiments.
We report the first finding of modern planktonic foraminifera in seafloor sediment samples collected from offshore Abu Dhabi, UAE. Samples containing planktonic foraminifera are found at water depths of >30 m in the northern sector of the UAE offshore. The planktonic assemblage is dominated by the minute triserial species Neogallitellia vivans, with sporadic occurrences of the species Globigerinoides tenellus, as well as Turborotalita cf. clarkei. Additionally, we report the occurrence of the meroplanktonic species Tretomphalus bulloides, possessing float chambers, and an enigmatic microfossil tentatively identified as Bachmayerella sp.
Detailed SEM imaging of the wall of N. vivans shows that it possesses a thin (1–2 µm) wall, as well as micropores (<0.5 µm) that often occur in doublets and triplets. We speculate that its pore arrangements may be an adaptation for life in low-oxygen conditions such as those found in eutrophic or upwelling areas in marginal seas.
The high relative abundance of N. vivans offshore Abu Dhabi suggests that it may be an opportunistic or invasive species in the Arabian Gulf. Future research utilizing plankton nets or sediment traps, as well as Holocene sediment cores from this region, may determine whether these foraminifera represent an active, living population in the present-day Arabian Gulf or are merely recent immigrants transported by currents or in ballast water.
Imaged specimens are currently deposited in the Micropaleontology Laboratory in College of Petroleum Engineering and Geosciences (CPG), King Fahd University of Petroleum and Minerals (KFUPM). The raw high-quality SEM specimens that we captured from our laboratory at KFUPM are available in the Supplement (Samples S1a–S5c). The samples will be transferred to the permanent archive at the European Micropalaeontological Reference Centre at Micropress Europe, Kraków, Poland.
The supplement related to this article is available online at https://doi.org/10.5194/jm-45-415-2026-supplement.
TS, with the support of his technical team, conducted the sampling campaign and providing SDP and MAK with the raw materials in the form of loose sediments and data on seawater temperature and salinity. SDP conducted the basic and advanced sample processing from raw materials, including sample preparation, laboratory analysis, data collection, and interpretation from the prepared materials, which progressed under the supervision of TS and MAK. SDP, with the support of MAK, conducted the taxonomical description and the addition of interrelated information. SDP, TS, and MAK, simultaneously compiled the literature review of the described species, including some materials written in German. The final paper was prepared by SDP, with equal contributions from all of the co-authors.
The contact author has declared that none of the authors has any competing interests.
Publisher's note: Copernicus Publications remains neutral with regard to jurisdictional claims made in the text, published maps, institutional affiliations, or any other geographical representation in this paper. The authors bear the ultimate responsibility for providing appropriate place names. Views expressed in the text are those of the authors and do not necessarily reflect the views of the publisher.
The Environment Agency Abu Dhabi (EAD) is gratefully acknowledged for facilitating the use of the research vessel Jaywun to collect sediment samples. Ahmed Abdelmaksoud, Sruthi Padampath Sreenivasan, and Salvador Rojas Lequerica are thanked for the collection and initial processing of the samples. We thank Syouma Hikmahtiar and Mohammed N. Al-Khabbaz for their technical support during the initial SEM imaging (first stage) for the miscellaneous species being reported from the Desktop SEM. We also acknowledge the technical support from Asmaa Korin and Ajibola Hakeem Okeyode in the detailed observation (pores and ultrastructural features) with the FE-SEM (second stage) for the Supplement. We appreciate Luke Mander as the editor-in-chief and Maria Rose Petrizzo as the handling editor of our submission in the Journal of Micropalaeontology. We also gratefully acknowledge two anonymous reviewers and Kirsty Edgar for their suggestions and commentaries that served to improve the paper. We express our gratitude to John W. Murray's family for their generosity in granting us the article processing cost (APC) waiver with regard to our submission to the special issue in this journal dedicated to honouring John W. Murray's monumental contributions towards our understanding of foraminiferal ecology.
SDP and MAK received support from the College of Petroleum Engineering and Geosciences, King Fahd University of Petroleum and Minerals. SDP received support from the Brian J. O'Neill Memorial Scholarship of the Grzybowski Foundation and the Kaminski Foundation for Micropaleontological Research. TS received support from the Khalifa University of Science and Technology through grant no. RIG-2023-061.
This paper was edited by Maria Rose Petrizzo and reviewed by Kirsty Edgar and two anonymous referees.
Alsharhan, A. S. and Kendall, C. G. St. C.: Holocene coastal carbonates and evaporites of the southern Arabian Gulf and their ancient analogues, Earth-Sci. Rev., 61, 191–243, https://doi.org/10.1016/S0012-8252(02)00110-1, 2003.
Alve, E.: A common opportunistic foraminiferal species as an indicator of rapidly changing conditions in a range of environments, Estuar. Coast. Shelf S., 57, 501–514, https://doi.org/10.1016/S0272-7714(02)00383-9, 2003.
Al-Yamani, F. Y., Skryabin, V., and Durvasula, S. R. V.: Suspected ballast water introductions in the Arabian Gulf, Aquat. Ecosyst. Health, 18, 282–289, https://doi.org/10.1080/14634988.2015.1027135, 2015.
Amao, A. O., Al-Ramadan, K., Kaminski, M., and Frontalini, F.: Benthic foraminiferal diversity in the Arabian Gulf: spatial patterns in a basin-wide assessment, PLoS ONE, 20, e0327033, https://doi.org/10.1371/journal.pone.0327033, 2025.
Banner, F. T., Pereira, C. P. G., and Desai, D.: “Tretomphaloid” float chambers in the Discorbidae and Cymbaloporidae, J. Foramin. Res., 15, 159–174, https://doi.org/10.2113/gsjfr.15.3.159, 1985.
Berger, W. H. and Diester-Haass, L.: Paleoproductivity: the benthic/planktonic ratio in foraminifera as a productivity index, Mar. Geol., 81, 15–25, https://doi.org/10.1016/0025-3227(88)90014-X, 1988.
Brock, J. C., McClain, C. R., Anderson, D. M., Prell, W. L., and Hay, W. W.: Southwest monsoon circulation and environments of recent planktonic foraminifera in the northwestern Arabian Sea, Paleoceanography, 7, 799–813, 1992.
Brummer, G.-J. A. and Kučera, M.: Taxonomic review of living planktonic foraminifera, J. Micropalaeontol., 41, 29–74, https://doi.org/10.5194/jm-41-29-2022, 2022.
Charlton, T. R., De Smet, M. E. M., Samodra, H., and Kaye, S. J.: The stratigraphic and structural evolution of the Tanimbar Islands, eastern Indonesia, J. Southe. Asian Earth, 6, 343–358, https://doi.org/10.1016/0743-9547(91)90080-H, 1991.
Cherif, O. H., Al-Rifaiy, I. A., and Al-Ghadban, A.-N.: Distribution of Foraminifera in the Arabian Gulf, Micropaleontology, 43, 253–280, https://doi.org/10.2307/1485827, 1997.
Cushman, J. A.: A Recent Gümbelitria (?) from the Pacific, Contrib. Cushman Lab. Foraminiferal Res., 10, 105, 1934.
Darling, K. F., Thomas, E., Kasemann, S. A., Seears, H. A., Smart, C. W., and Wade, C. M.: Surviving mass extinction by bridging the benthic/planktic divide, P. Natl. Acad. Sci. USA, 106, 12629–12633, https://doi.org/10.1073/pnas.0902827106, 2009.
De Smet, M. E. M., Fortuin, A. R., Troelstra, S. R., Van Marle, L. J., Karmini, M., Tjokrosapoetro, S., and Hadiwasastra, S.: Detection of collision-related vertical movements in the Outer Banda Arc (Timor, Indonesia), using micropaleontological data, J. Southe. Asian Earth, 4, 337–356, https://doi.org/10.1016/0743-9547(90)90005-X, 1990.
d'Orbigny, A. D.: Foraminifères, in: Histoire physique, politique et naturelle de l'ile de Cuba, edited by: de la Sagra, R., A. Bertrand, Paris, 1–224, 1839.
Ghosh, A.: Estuarine foraminifera from the Gulf of Cambay, J. Geol. Soc. India, 80, 65–74, https://doi.org/10.1007/s12594-012-0119-7, 2012.
Ghosh, A., Saha, S., Saraswati, P. K., Banerjee, S., Burley, S., and Rao, T. G.: Gallitellia – a proxy for palaeomonsoonal upwelling on the western coast of India?, Curr. Sci., 95, 1608–1611, 2008.
Ghosh, A., Saha, S., Saraswati, P. K., Banerjee, S., and Burley, S.: Intertidal foraminifera in the macro-tidal estuaries of the Gulf of Cambay: Implications for interpreting sea-level change in palaeo-estuaries, Mar. Petrol. Geol., 26, 1592–1599, https://doi.org/10.1016/j.marpetgeo.2008.08.002, 2009.
Glock, N., Schönfeld, J., and Mallon, J.: The Functionality of Pores in Benthic Foraminifera in View of Bottom Water Oxygenation: A Review, in: Anoxia: Evidence for Eukaryote Survival and Paleontological Strategies, Cellular Origin, Life in Extreme Habitats and Astrobiology, Vol. 21, edited by: Altenbach, A. V., Bernhard, J. M., and Seckbach, J., Springer, Dordrecht, 540–556, https://doi.org/10.1007/978-94-007-1896-8_28, 2012.
Gull, H. M., Tawabini, B. S., Amao, A. O., Prayudi, S. D., Ayranci, K., and Kaminski, M. A.: Benthic foraminiferal response to a salinity gradient in the Uqair hypersaline lagoonal system, Gulf Coast of Saudi Arabia, Micropaleontology, 71, 261–279, https://doi.org/10.47894/mpal.71.3.04, 2025.
Hamza, W. and Munawar, M.: Protecting and managing the Arabian Gulf: Past, present and future, Aquat. Ecosyst. Health, 12, 429–439, https://doi.org/10.1080/14634980903361580, 2009.
Hemleben, C.: Zur Morphogenese planktonischer Foraminiferen, Zitteliana, 1, 91–133, 1969.
Hemleben, C., Spindler, M., and Anderson, O. R.: Modern Planktonic Foraminifera, Springer, New York, 363 pp., https://doi.org/10.1007/978-1-4612-3544-6, 1989.
Hofker, J.: Tertiary foraminifera of coastal Ecuador- Part II Additional notes on the Eocene species, J. Paleontol., 30, 891–958, 1956.
Hughes Clarke, M. W. and Keij, A. J.: Organisms as producers of carbonate sediment and indicators of environment in the southern Persian Gulf, in: The Persian Gulf, edited by: Purser, B. H., Springer-Verlag, Berlin, 33–56, https://doi.org/10.1007/978-3-642-65545-6_3, 1973.
Ivanova, E., Schiebel, R., Singh, A. D., Schmiedl, G., Niebler, H.-S., and Hemleben, C.: Primary production in the Arabian Sea during the last 135 000 years, Palaeogeogr. Palaeocl., 197, 61–82, https://doi.org/10.1016/S0031-0182(03)00386-9, 2003.
Kassler, P.: The structural and geomorphic evolution of the Persian Gulf, in: The Persian Gulf, edited by: Purser, B. H., Springer-Verlag, Berlin, 11–32, https://doi.org/10.1007/978-3-642-65545-6_2, 1973.
Katz, M. E., Cramer, B. S., Franzese, A., Hönisch, B., Miller, K. G., Rosenthal, Y., and Wright, J. D.: Traditional and emerging geochemical proxies in foraminifera, J. Foramin. Res., 40, 165–192, https://doi.org/10.2113/gsjfr.40.2.165, 2010.
Keller, G.: Biotic effects of impacts and volcanism, Earth Planet. Sc. Lett., 215, 249–264, https://doi.org/10.1016/S0012-821X(03)00390-X, 2003.
Keller, G. and Pardo, A.: Disaster opportunists Guembelitrinidae: Index for environmental catastrophes, Mar. Micropaleontol., 53, 83–116, https://doi.org/10.1016/j.marmicro.2004.04.012, 2004.
Kimoto, K., Ishimura, T., Tsunogai, U., Itaki, T., and Ujiié, Y.: The living triserial planktic foraminifer Gallitellia vivans (Cushman): distribution, stable isotopes, and paleoecological implications, Mar. Micropaleontol., 71, 71–79, https://doi.org/10.1016/j.marmicro.2009.01.006, 2009.
Kranner, M., Harzhauser, M., Mandic, O., Strauss, P., Siedl, W., and Piller, W. E.: Trends in temperature, salinity and productivity in the Vienna Basin (Austria) during the early and middle Miocene, based on foraminiferal ecology, Palaeogeogr. Palaeocl., 581, 110640, https://doi.org/10.1016/j.palaeo.2021.110640, 2021.
Kroon, D. and Nederbragt, A. J.: Ecology and paleoecology of triserial planktic foraminifera, Mar. Micropaleontol., 16, 25–38, https://doi.org/10.1016/0377-8398(90)90027-J, 1990.
Lambeck, K.: Shoreline reconstructions for the Persian Gulf since the last glacial maximum, Earth Planet. Sc. Lett., 142, 43–57, https://doi.org/10.1016/0012-821X(96)00069-6, 1996.
Liu, H., Nour El-Din, N., Rowe, G. T., Al-Ansi, M., Wei, C.-L., Soliman, Y., Nunnally, C. C., Quigg, A., Al-Ansari, I. S., Al-Maslamani, I., and Abdel-Moati, M. A.: Characteristics and renewal of zooplankton communities under extreme environmental stresses in the oligotrophic hypersaline Arabian Gulf, Prog. Oceanogr., 201, 102643, https://doi.org/10.1016/j.pocean.2021.102643, 2022.
Loeblich Jr., A. R. and Tappan, H.: Some new and revised genera and families of hyaline calcareous Foraminiferida (Protozoa), Trans. Am. Microscop. Soc., 239–265, https://doi.org/10.2307/3226297, 1986.
Loeblich, A. R. and Tappan, H.: Foraminiferal Genera and their Classification, Van Nostrand Reinhold, New York, 970 pp., https://doi.org/10.1007/978-1-4899-5760-3, 1987.
Lutze, G. F.: Benthische Foraminiferen in Oberflächen-Sedimenten des Persischen Golfes, Teil 1: Arten, Meteor Forschungsergeb. Reihe C, 17, 1–66, 1974.
Naidu, P. D.: Distribution of upwelling index planktonic-foraminifera in the sediments of the western continental-margin of India, Oceanol. Acta, 13, 327–333, 1990.
Özdikmen, H.: Substitute names for some unicellular animal taxa (Protozoa), Munis Entomol. Zool., 4, 233–256, 2009.
Pardo, A. and Keller, G.: Biotic effects of environmental catastrophes at the end of the Cretaceous and early Tertiary: Guembelitria and Heterohelix blooms, Cretac. Res., 29, 1058–1073, https://doi.org/10.1016/j.cretres.2008.05.031, 2008.
Parker, F. L.: Eastern Mediterranean Foraminifera, Reports of the Swedish Deep-Sea Expedition, Sediment Cores from the Mediterranean Sea and the Red Sea, 8, 217–283, 1958.
Pawlowski, J.: Distribution and taxonomy of some benthic tiny foraminifers from the Bermuda Rise, Micropaleontology, 37, 163–172, 1991.
Peeters, F. J. C., Ivanova, E., Conan, S. M. H., Brummer, G. J. A., Ganssen, G. M., Troelstra, S. R., and van Hinte, J. E.: Size analysis of planktic foraminifera from the Arabian Sea, Mar. Micropaleontol., 36, 31–63, https://doi.org/10.1016/S0377-8398(98)00026-7, 1999.
Peryt, D., Gedl, P., Worobiec, E., Worobiec, G.. and Peryt, T.M.: Foraminiferal and palynological records of an abrupt environmental change at the Badenian/Sarmatian boundary (Middle Miocene): a case study in northeastern Central Paratethys, Geosciences, 14, 86, https://doi.org/10.3390/geosciences14030086, 2024.
Prayudi, S. D., Tawabini, B. S., Korin, A., and Kaminski, M. A.: Morphological deformities in benthic foraminiferal assemblages from shallow marine and lagoonal environments of the western Arabian Gulf, J. Foraminifer. Res., 54, 290–308, https://doi.org/10.61551/gsjfr.54.3.290, 2024a.
Prayudi, S. D., Tawabini, B. S., Amao, A. O., Korin, A., Gull, H. M., Arrofi, D., and Kaminski, M. A.: Survival of biocalcifying shallow-marine benthic organisms in the coastal areas of the Arabian Gulf under conditions of global warming: Is there a limit to their resilience?, Palaeogeogr. Palaeocl., 653, 112423, https://doi.org/10.1016/j.palaeo.2024.112423, 2024b.
Prayudi, S., Steuber, T., and Kaminski, M. A.: Agglutinated benthic foraminifera from the southeastern Arabian Gulf: A taxonomical and ecological perspective, Micropaleontology, 72, 361–380, 2026.
Purser, B. H. and Seibold, E.: The principal environmental factors influencing Holocene sedimentation and diagenesis in the Persian Gulf, in: The Persian Gulf: Holocene Carbonate Sedimentation and Diagenesis in a Shallow Epicontinental Sea, edited by: Purser, B. H., Springer-Verlag, Berlin, 1–10, https://doi.org/10.1007/978-3-642-65545-6_1, 1973.
Rögl, F. and Franz, H. E.: Bachmayerella – ein neues problematisches Mikrofossil aus dem marinen Mittelmiozän von Walbersdorf, Burgenland, Ann. Naturhist. Mus. Wien, 82, 83–98, 1979.
Reynolds, R. M.: Physical oceanography of the Gulf, Strait of Hormuz, and the Gulf of Oman – results from the Mt. Mitchell expedition, Mar. Pollut. Bull., 27, 35–59, 1993.
Rögl, F. and Bolli, H. M.: Holocene to Pleistocene Planktonic Foraminifera of Leg 15, Site 147 (Cariaco Basin [Trench], Caribbean Sea) and Their Climatic Interpretation, Initial Reports of the Deep Sea Drilling Project, 15, 553–615, https://doi.org/10.2973/dsdp.proc.15.113.1973, 1973.
Schenk, B., Gebhardt, H., Wolfgring, E., and Zorn, I.: Cyclic paleo-salinity changes inferred from benthic foraminiferal assemblages in the Upper Burdigalian (Lower Miocene) Korneuburg Basin, Austria, Palaeogeogr. Palaeocl., 490, 473–487, https://doi.org/10.1016/j.palaeo.2017.11.027, 2018.
Schiebel, R. and Hemleben, C.: Planktic foraminifers in the modern ocean, 358, Springer, Berlin, https://doi.org/10.1007/978-3-662-50297-6, 2017.
Schiebel, R., Waniek, J., Bork, M., and Hemleben, C.: Planktic foraminiferal production stimulated by chlorophyll redistribution and entrainment of nutrients, Deep-Sea Res. Pt. I, 48, 721–740, https://doi.org/10.1016/S0967-0637(00)00065-0, 2001.
Schiebel, R., Zeltner, A., Treppke, U. F., Waniek, J. J., Bollmann, J., Rixen, T., and Hemleben, C.: Distribution of diatoms, coccolithophores and planktic foraminifers along a trophic gradient during SW monsoon in the Arabian Sea, Mar. Micropaleontol., 51, 345–371, 2004.
Schönfeld, J., Alve, E., Geslin, E., Jorissen, F., Korsun, S., Spezzaferri, S., and Members of the FOBIMO group: The FOBIMO (FOraminiferal BIo-MOnitoring) initiative – Towards a standardised protocol for soft-bottom benthic foraminiferal monitoring studies, Mar. Micropaleontol., 94–95, 1–13, https://doi.org/10.1016/j.marmicro.2012.06.001, 2012.
Schulz, H., von Rad, U., and Ittekkot, V.: Planktic foraminifera, particle flux and oceanic productivity off Pakistan, NE Arabian Sea: modern analogues and application to the palaeoclimatic record, Geol. Soc. Lond. Spec. Publ., 195, 499–516, https://doi.org/10.1144/GSL.SP.2002.195.01.27, 2002.
Sheppard, C., Price, A., and Roberts, C.: Marine Ecology of the Arabian Region: Patterns and Processes in Extreme Tropical Environments, Academic Press, London, 359 pp., ISBN 9780126394900, 1992.
Sheppard, C., Al-Husiani, M., Al-Jamali, F., Al-Yamani, F., Baldwin, R., Bishop, J., Benzoni, F., Dutrieux, E., Dulvy, N. K., Durvasula, S. R. V., Jones, D. A., Loughland, R., Medio, D., Nithyanandan, M., Pilling, G. M., Polikarpov, I., Price, A. R. G., Purkis, S., Riegl, B., Saburova, M., Samimi-Namin, K., Taylor, O., Wilson, S., and Zainal, K.: The Gulf: A young sea in decline, Mar. Pollut. Bull., 60, 13–38, https://doi.org/10.1016/j.marpolbul.2009.10.017, 2010.
Spezzaferri, S. and Rögl, F.: Bolboforma (Phytoplankton Incertae Sedis), Bachmayerella and other Calciodinelloidea (Phytoplankton) from the Middle Miocene of the Alpine–Carpathian Foredeep (Central Paratethys), J. Micropalaeontol., 23, 139–152, https://doi.org/10.1144/jm.23.2.139, 2004.
Steuber, T., Sreenivasan, S. P., Rojas Lequerica, S., Abdelmaksoud, A., Al Jaberi, A., Al Shehhi, A., Prayudi, S., Kaminski, M. A., and Al-Suwaidi, A.: Modern carbonate sedimentation in an extreme marine environment: the southeastern Arabian Gulf, Mar. Geol., 496, 107764, https://doi.org/10.1016/j.margeo.2026.107764, 2026.
Subba Rao, D. V. and Al-Yamani, F.: Phytoplankton ecology in the waters between Shatt Al-Arab and Straits of Hormuz, Arabian Gulf: A review, Plankton Biol. Ecol., 45, 101–116, 1998.
Ujiié, Y., Kimoto, K., and Pawlowski, J.: Molecular evidence for an independent origin of modern triserial planktonic foraminifera from benthic ancestors, Mar. Micropaleontol., 69, 334–340, https://doi.org/10.1016/j.marmicro.2008.09.003, 2008.
Van der Zwaan, G. J., Jorissen, F. J., and De Stigter, H. C.: The depth dependency of planktonic/benthic foraminiferal ratios: constraints and applications, Mar. Geol., 95, 1–16, https://doi.org/10.1016/0025-3227(90)90016-D, 1990.
Vasou, P., Vervatis, V., Krokos, G., Hoteit, I., and Sofianos, S.: Variability of water exchanges through the Strait of Hormuz, Ocean Dynam., 70, 1053–1065, https://doi.org/10.1007/s10236-020-01384-2, 2020.
Versteegh, G. J. M., Servais, T., Streng, M., Munnecke, A., and Vachard, D.: A discussion and proposal concerning the use of the term calcispheres, Palaeontology, 52, 343–348, https://doi.org/10.1111/j.1475-4983.2009.00854.x, 2009.
Williamson, W. C.: On the Recent Foraminifera of Great Britain, Ray Society, London, 107 pp., ISBN 9780341764762, 1858.
Wyville-Thomson, C.: The Voyage of the “Challenger”: The Atlantic. A preliminary account of the general results of the exploring voyage of H.M.S. “Challenger” during the year 1873 and the early part of the year 1876, Vol. 1, Macmillan and Co., London, 424 pp., https://doi.org/10.5962/bhl.title.79255, 1878.
Zheng, S. and Fu, Z.: Class Foraminifera, in: Checklist of Marine Biota of China Seas, edited by: Liu, J. Y., China Science Press, Beijing, 108–174, ISBN 9787030237224, 2008.
Building on historical research, we report the discovery of planktonic foraminifera in sediments offshore Abu Dhabi. Surprisingly, the samples are dominated by Neogallitellia vivans, a rare species typically found in the Indo-Pacific. Our detailed analysis of its shell structure suggests that this tiny organism is uniquely adapted to the extreme salinity of the southern Arabian Gulf. This finding highlights the ability of marine life to thrive in harsh, hypersaline environments.
Building on historical research, we report the discovery of planktonic foraminifera in sediments...