the Creative Commons Attribution 4.0 License.
the Creative Commons Attribution 4.0 License.
 | 03 Jun 2026
| 03 Jun 2026
Ontogenetic growth of three cultured species of Heterocypris Claus, 1892 (Crustacea: Ostracoda): eggs and valve morphology
Mauricio Bonilla-Flores
Liseth Pérez
Peter Frenzel
Paula Echeverría-Galindo
Junbo Wang
Antje Schwalb
Ontogenetic information remains limited for many cypridoid ostracods despite its importance for reliable taxonomy, species identification, and the interpretation of ecological patterns based on size and morphology. This is particularly relevant in paleoecological studies, where, usually, only valves are preserved. We investigated post-embryonic development in three asexual species of Heterocypris inhabiting ephemeral aquatic environments: Heterocypris exodonta from the Tibetan Plateau, Heterocypris incongruens from Mexico, and Heterocypris salina from Germany. Based on laboratory cultures maintained under controlled conditions, egg morphology, juvenile valve development, and ontogenetic growth were quantified using measurements of valve length and height. All three species exhibited nine developmental stages, comprising eight juvenile instars and one adult stage. Eggs possess a rough external surface and a distinct internal eggshell structure. Early juvenile stages display a marked polygonal reticulation pattern on their valves that progressively weakens and disappears in adults. Growth ratios, defined as the proportional increase in valve size between successive developmental stages, averaged values close to those expected under geometric growth. However, considerable variation is reported among species and developmental stages. The results provide reference data on ontogenetic development and diagnostic morphological characters, improving the distinction between juvenile stages and both intraspecific and interspecific variation and supporting the interpretation of modern and fossil assemblages.
- Article
(16336 KB) - Full-text XML
-
Supplement
(385 KB) - BibTeX
- EndNote




Non-marine ostracods are bivalved crustaceans with soft parts enclosed within a carapace composed of a right and a left valve (Boomer et al., 2003). These arthropods grow by a specific number of molts, and ontogeny comprises the egg, juvenile instars, and adult stages, which, together, ensure completion of the life cycle and population persistence (Meisch, 2000; Dumont et al., 2002; Pereira et al., 2017).
Ostracod eggs are surrounded by a multilayered eggshell formed during oogenesis. Classical histological studies indicate that this structure comprises at least two principal layers: an inner layer produced by the oocyte and an outer layer secreted by the ovarian epithelium during oogenesis (Woltereck, 1898; Weygoldt, 1960; Hartmann, 1968; Roessler, 1982). Furthermore, Kesling (1951) described the eggshell as a double-walled structure composed of chitin impregnated with calcium carbonate, with the inner wall separated from the outer wall and connected to it by fine chitinous processes, indicating a complex eggshell organization before hatching. In addition, SEM studies have documented external egg morphology, particularly in Heterocypris incongruens, including surface ornamentation (Dumont et al., 2002; Özuluğ and Suludere, 2012), which may contribute to protection against adverse environmental conditions such as drying and extreme temperatures (Smith et al., 2015).
Resting (dormant) eggs play a key ecological role in non-marine ostracods by enabling populations to persist under fluctuating or adverse environmental conditions (Rossi et al., 2012). This leads to the formation of egg banks, defined as the accumulation of viable dormant eggs in the sediment that hatch when conditions become favorable (De Stasio, 2007; Hairston and Fox, 2009; Rosa et al., 2020, 2023). Egg banks act as temporal reservoirs that enhance resilience in temporary aquatic systems and provide insights into the evolutionary history and adaptive capabilities of these microcrustaceans (Bellin et al., 2020; Wang et al., 2025).
Ontogeny in non-marine ostracods is fundamental to taxonomy and classification as developmental changes in valve morphology are key for species identification and interpreting phylogenetic relationships in modern and fossil assemblages (De Deckker and Martens, 2013). Most podocopid ostracods, particularly within the Cyprididae, exhibit nine post-embryonic instars, including eight juvenile stages and one adult stage (Smith and Martens, 2000). However, in some Heterocypris species, such as H. bogotensis Roessler, 1982, H. bosniaca Petkowski et al. (2000) (Aguilar-Alberola and Mesquita-Joanes, 2013), and H. incongruens (Rossi et al., 2015), an additional early developmental stage (A-9), corresponding to a prenaupliar stage, has been reported, resulting in a total of 10 instars. Nonetheless, juvenile morphology remains poorly documented despite its importance for avoiding taxonomic misidentifications. Valve characteristics often allow straightforward identification of adult ostracods. In most cases, identification of juveniles at the species level is considerably more challenging, even when soft-part morphology is considered (Holmes, 2001; Lajblová et al., 2014). In parallel, the size, shape, ornamentation, and scar pattern of valves are primarily used for identification. However, ecophenotypic variation, observable morphological differences within a population resulting from environmental influences, should be considered as well (Holmes, 2001). Mainly, water temperature, conductivity, and food availability strongly influence ostracod ontogeny, driving spatial and temporal variations in size and shape across populations, including seasonal and interannual fluctuations (Baltanás et al., 2000).
Recognizing modern analog species is crucial for identifying fossil material at the species level and highlights the importance of ostracod cultures for understanding all developmental instars (Boomer et al., 2003). Under natural conditions, first juveniles generally preserve poorly, mainly because of weak calcification and fragile valves (Tinn and Meidla, 2003). Also, this may be further underrepresented in sediments due to cannibalism by adults or late instars (A-1) (Schreiber, 1922, and personal observation). Nevertheless, lake sediment provides valuable records for studying ontogeny through preserved valves, allowing reconstruction of evolutionary processes and environmental changes, as well as distinguishing autochthonous from allochthonous assemblages (Baltanás et al., 2000; Boomer et al., 2003).
To distinguish juveniles from adults, changes reflecting the addition of new appendages and, in sexual species, the onset of sexual maturity can be used as diagnostic criteria (Retrum and Kaesler, 2005). In males, hemipenis morphology is taxonomically important for identification because their shape differs among species. Meanwhile, in asexual populations, lacking males, the examination of the female genital lobes (vaginas or genital openings) holds significant potential for distinguishing morphologically similar species. This is reflected in interspecific variation in the intersection (junction of seminal ducts or oviduct), which corresponds to the chitinous frame of the vaginal opening sensu Kesling (1951) (see Fontana and Ballent, 2005; Karanovic and Lee, 2012; Kong et al., 2014; Matzke-Karasz et al., 2017; Mesquita-Joanes et al., 2020). This distinction can be of great utility when the morphology of valves and anatomical appendages shows limited interspecific variation (Bonilla-Flores et al. 2024, 2025).
Growth in ostracods is often described by Brooks' rule, which suggests that crustaceans' size doubles their volume through each molt, and this increase shows a linear relationship roughly up to the square root of 2, resulting in the coefficient of 1.26 with each molt (Brooks, 1886). However, deviations in non-marine ostracods are common (Hartmann, 1968; Baltanás et al., 2000; Mezquita et al., 2000, 2002; Van Doninck et al., 2003; Danielopol et al., 2008; Zhai et al., 2015; Mao et al., 2021), limiting its taxonomic value (Martens, 1985; Watabe and Kaesler, 2004), although it remains useful for assessing ontogenetic variability and environmental influences in fossil assemblages (McCormack et al., 2019).
Genus Heterocypris is one of the most diverse within the Cyprididae family, with 72 species (Meisch et al., 2024), and detailed comparative ontogenetic studies remain scarce. In particular, SEM-based analyses of valve morphology across instars are still limited, even for widely distributed species such as H. incongruens and H. salina. This limitation contributes to longstanding uncertainty regarding whether their broad geographic distributions reflect truly cosmopolitan species or complexes of cryptic taxa (Yoo et al., 2017). Recent molecular and population genetic studies indicate substantial genetic structuring and the presence of multiple clonal lineages in these taxa (Rossi et al., 2003, 2006) and, in some cases, suggest the existence of cryptic species complexes (Bonilla-Flores et al., 2025; Kilikowska et al., 2024), but ontogenetic data remain scarce. This underscores the need for integrative approaches combining ontogenetic and SEM-based morphological analyses. To address this gap, this study examines three species from different regions: H. exodonta from Nam Co, Tibetan Plateau (TP) (Bonilla-Flores et al., 2025); H. salina from Braunschweig, Germany; and H. incongruens from San Nicolás Tetelco, Mexico City. These species were selected based on their morphological similarity, occurrence in ephemeral environments, and suitability for laboratory culture. The objectives of this study are to (1) describe valve ontogeny, (2) characterize egg morphology, and (3) evaluate growth ratios to identify potential differences in developmental patterns.
The individuals of H. exodonta were collected in the study area located at 4728 m above sea level (m a.s.l.) from a temporary pond near Nam Co, a lake on the southern Tibetan Plateau (Table 1). Such temporary pools and lagoons formed around the lake because of wave action and the uneven terrain along the shoreline (Echeverría-Galindo, personal observation). This endorheic lake is highly exposed to solar radiation, the Indian Summer Monsoon, and the Westerlies (Zhu et al., 2008). Winters on the south-central Tibetan Plateau are dry, with precipitation mainly occurring between May and September (Anslan et al., 2020). Additionally, individuals of H. incongruens were collected from a flowerpot in San Nicolás Tetelco, Milpa Alta, southern Mexico City. The pots were watered with tap water and, particularly between June and August, with rainwater. The rainy season extends from June to September, while the dry season occurs during the rest of the year (Bouvier et al., 1993). Furthermore, specimens of H. salina were collected at the Botanical Garden in Braunschweig, Germany, an urban green area characterized by semi-controlled environmental conditions. The study site consisted of flowerpots exposed to natural climatic conditions, primarily irrigated with tap water. During the summer months, corresponding to the local rainy season (June–September), irrigation was supplemented by rainfall (Table 1). All ostracods were collected using a spatula and a hand net with a mesh size of 125 µm.
3.1 Laboratory work
The cultures of H. exodonta, H. incongruens, and H. salina were maintained under the same laboratory temperature (18–23 °C) and a photoperiod of 14:10 h of light to ensure comparability. Under these conditions, the species showed a relatively short cycle between hatching and the final moult of approximately 2 to 3 months. Temperature may influence growth rates and developmental timing; therefore, these values should be considered to be specific to the experimental conditions. The three species were fed spinach, and the cultures were maintained in Vilsa naturelle water (VILSA-BRUNNEN Otto Rodekohr GmbH); its parameters are given in Table 2.
Table 2Physico-chemical water characteristics of the water used for ostracod cultures.

A multiwell cell culture dish (Nunc™) with six compartments, each containing 10 mL of water, was used. When the juveniles were separated, each was fed with small pieces of spinach, and the water was renewed every 3 d, reducing the effects of bacteria or fungi growth.
The purpose of keeping living ostracods was to produce viable cultures that would reproduce multiple times to obtain enough valves for subsequent analysis. It was observed that, during initial trials, juvenile A-8, separated from the group, did not survive despite being provided with spinach as a substrate for feeding. Individually, they did not survive beyond 7–9 d after hatching. However, when multiple juveniles (10–15) were introduced into the same compartment, it was observed that the ostracods had the possibility of surviving for more days (25–30) and reaching adulthood. This is likely due to their tendency to aggregate when water conditions and access to food are favorable (personal observation). Additionally, solitary juveniles showed a higher susceptibility to fungal contamination than individuals maintained in aggregation. Nevertheless, the survival of some solitary specimens was observed after transfer to a clean container and thorough cleaning of the multiwell plates.
Maintaining healthy cultures was also essential for evaluating egg resistance to desiccation. To this end, eggs of H. exodonta were exposed to dry conditions for 120 h. Eggs were then rehydrated with distilled water, and morphological changes were monitored during rehydration.
The valves were separated using a size-0 brush from the culture dish and were stored in micropaleontological slides. Subsequently, they were photographed by scanning electron microscopy (SEM, ZEISS EVO Ls 25) at the Institute for Chemical and Thermal Process Engineering, Faculty of Mechanical Engineering, TU Braunschweig. The biological material is temporarily stored at the Institute of Geosystems and Bioindication, TU Braunschweig.
3.2 Statistical analysis
The length and height of individual valves and carapaces of adults and juveniles were measured using a Leica M125 compound microscope. Data were analyzed and plotted in R Studio using the ggplot2 package (R Core Team, 2022).
The mean growth ratio was calculated using valve measurements from juvenile and adult instars. The ratio relates the average increments in length and height from one instar to the following growth stage. To estimate the average growth ratio, individual ratios were summed and divided by the number of growth phases. Values close to those expected under geometric growth (∼ 1.260) were considered to be consistent with Brooks' rule, although deviations are expected due to ontogenetic differences among instars, interspecific and intraspecific variation, and environmental influences such as temperature (Brooks, 1886; Watabe and Kaesler, 2004). The systematic position of suprageneric taxa follows Meisch et al. (2024).
4.1 Systematic list
-
Class Ostracoda Latreille, 1802
-
Subclass Podocopa Sars, 1866
-
Order Podocopida Sars, 1866
-
Suborder Cypridocopina Baird, 1845
-
Superfamily Cypridoidea Baird, 1845
-
Subfamily Cyprinotinae Bronstein, 1947
-
Genus Heterocypris Claus, 1892
-
Heterocypris exodonta Bonilla-Flores and Karanovic in Bonilla-Flores et al. (2025)
Material. Females from a pond near the Nam Co Monitoring and Research Station for Multisphere Interactions (NAMORS), Institute of Tibetan Plateau Research, Chinese Academy of Sciences.
Reproduction. Asexual.
Ontogeny. Egg, eight juvenile instars, and one adult stage (Fig. 1).
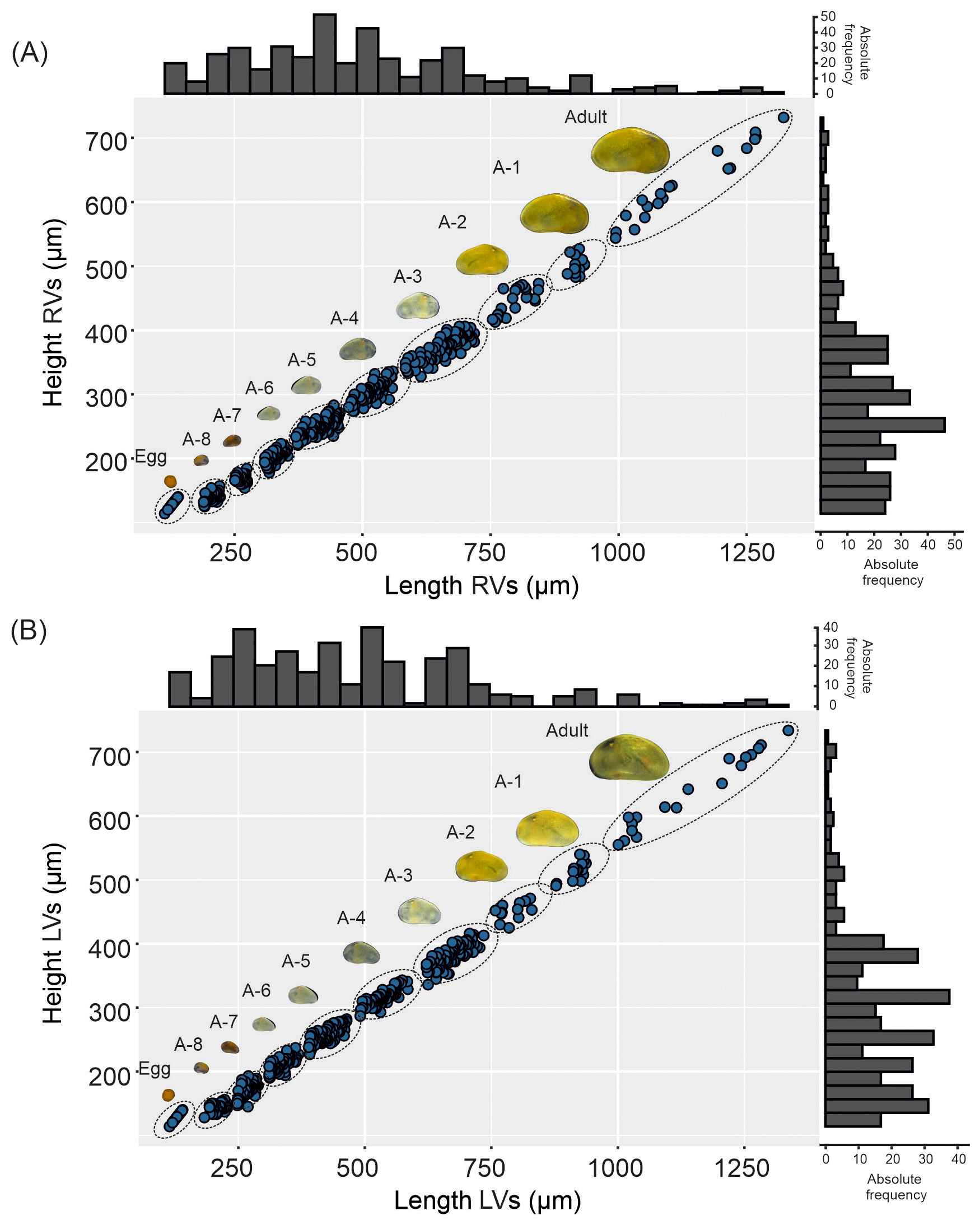
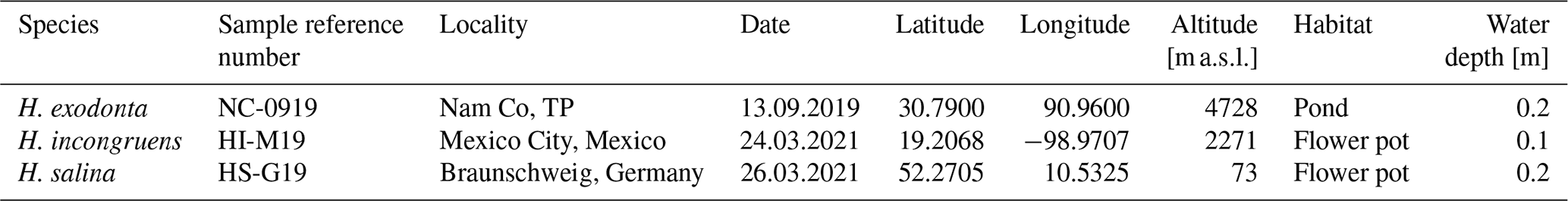
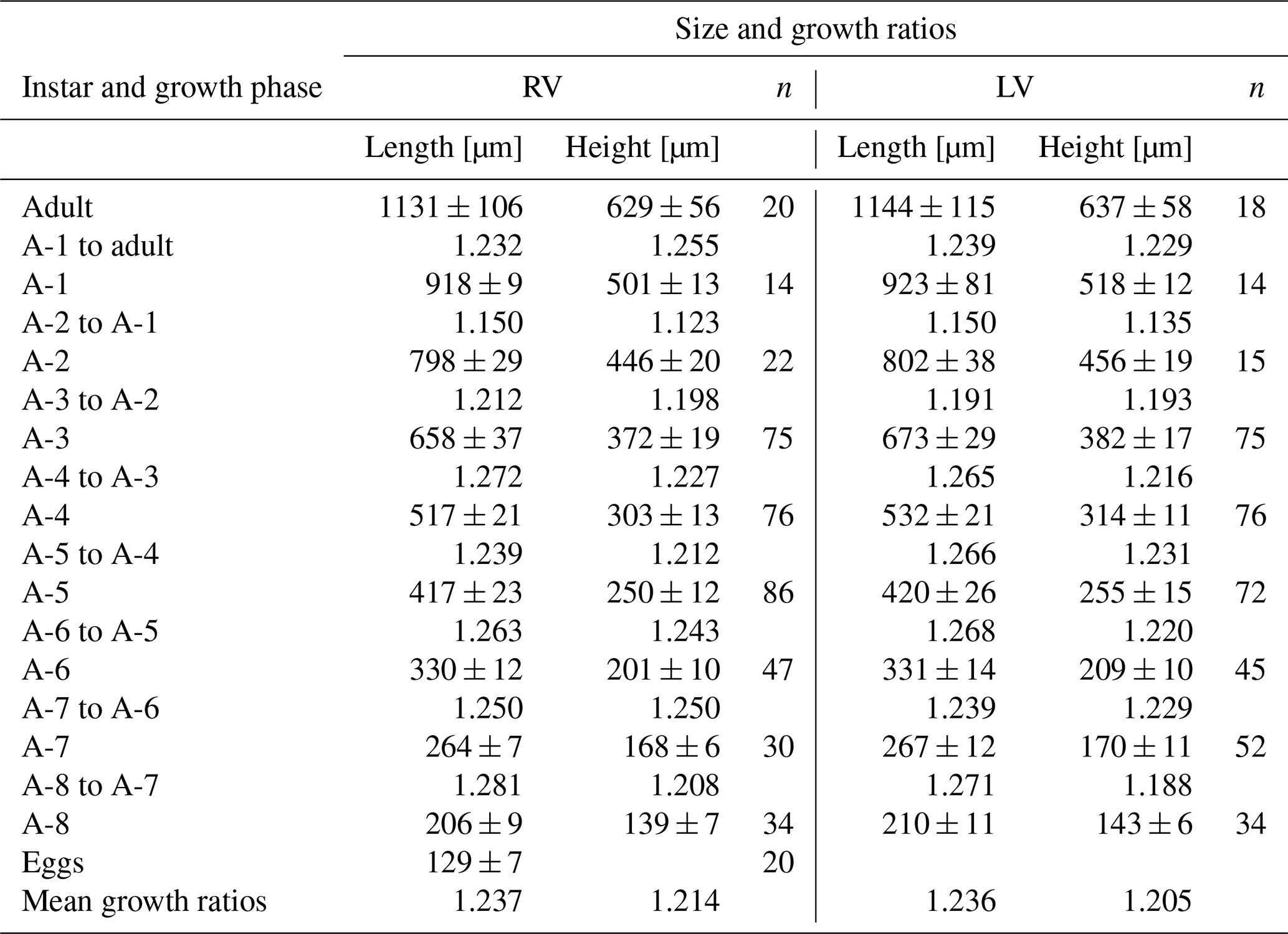
Figure 1Valve ontogeny of Heterocypris exodonta; from Nam Co, Tibetan Plateau (NC-0919). The scatterplot shows the measures of the length and height of (A) right valves (RVs), n=424, and (B) left valves (LVs), n=430. The histograms display the abundances of eggs, juvenile instars, and adult stage.
Egg characteristics. The eggs display an orange to yellowish coloration (Fig. 2A). The outer surface appears to be rough and is frequently covered by adhering particles such as sediment, diatoms, or plant fragments (Fig. 2B). These particles are likely attached by adhesive secretions during oviposition and do not constitute structural components of the eggshell (Wohlgemuth, 1914). The eggshell itself consists of two layers. The outer surface may appear to be porous or irregular, which could result from post-depositional changes rather than representing the primary structure. After stage-A-8 hatching, the eggshell shows an irregular rupture rather than a predefined opening (Fig. 2E), indicating that no distinct operculum is present (Hartmann, 1968). When dried (120 h), the eggs tended to collapse (Fig. 2F), but after rehydration they turned orange after 20 min and regained their original diameter after 60 min. The average diameter of the eggs is 129 µm.

Figure 2Heterocypris exodonta. (A) Egg cluster adhered to a plant fragment; (B) egg showing adhered particles on the surface and an opening with exposed eggshell layers; (C–D) higher-magnification views of the eggshell at the opening, illustrating the inner and outer eggshell layers; (E) irregular opening after hatching; and (F) collapsed egg after desiccation.
Adult valves. The coloration is yellowish (see Fig. 1). The external surface has numerous normal pores, each with a single seta. With a lateral view, a small hump is noticeable on the dorsal side. The RV features a broad inner lamella with distinct tubercles on both the anterior and posterior margins. The left valve (LV) is taller and longer, overlapping the right valve (RV).
Juvenile valves. External surface with normal pores; reticulation on the surface of the valves is observable in the early instars, from A-8 to A-2 (Figs. 3 and 4), and gradually dissipates in later instars. Stage A-8 is characterized by a rounded shape, which then develops into a more triangular form in the subsequent molting instars (A-7 to A-2) (Fig. 3). Stage A-8 shows RVs with 206 µm in length and 139 µm in height, on average, and LVs measuring 210 µm in length and 143 µm in height. In contrast, adults display RVs averaging 1131 µm in length and 629 µm in height, and LVs have an average length of 1144 µm and a height of 637 µm (Table 2). The mean growth ratio (length height) for the RV is 1.237 and 1.214, and for the LV it is 1.236 and 1.205, respectively (Table 2).
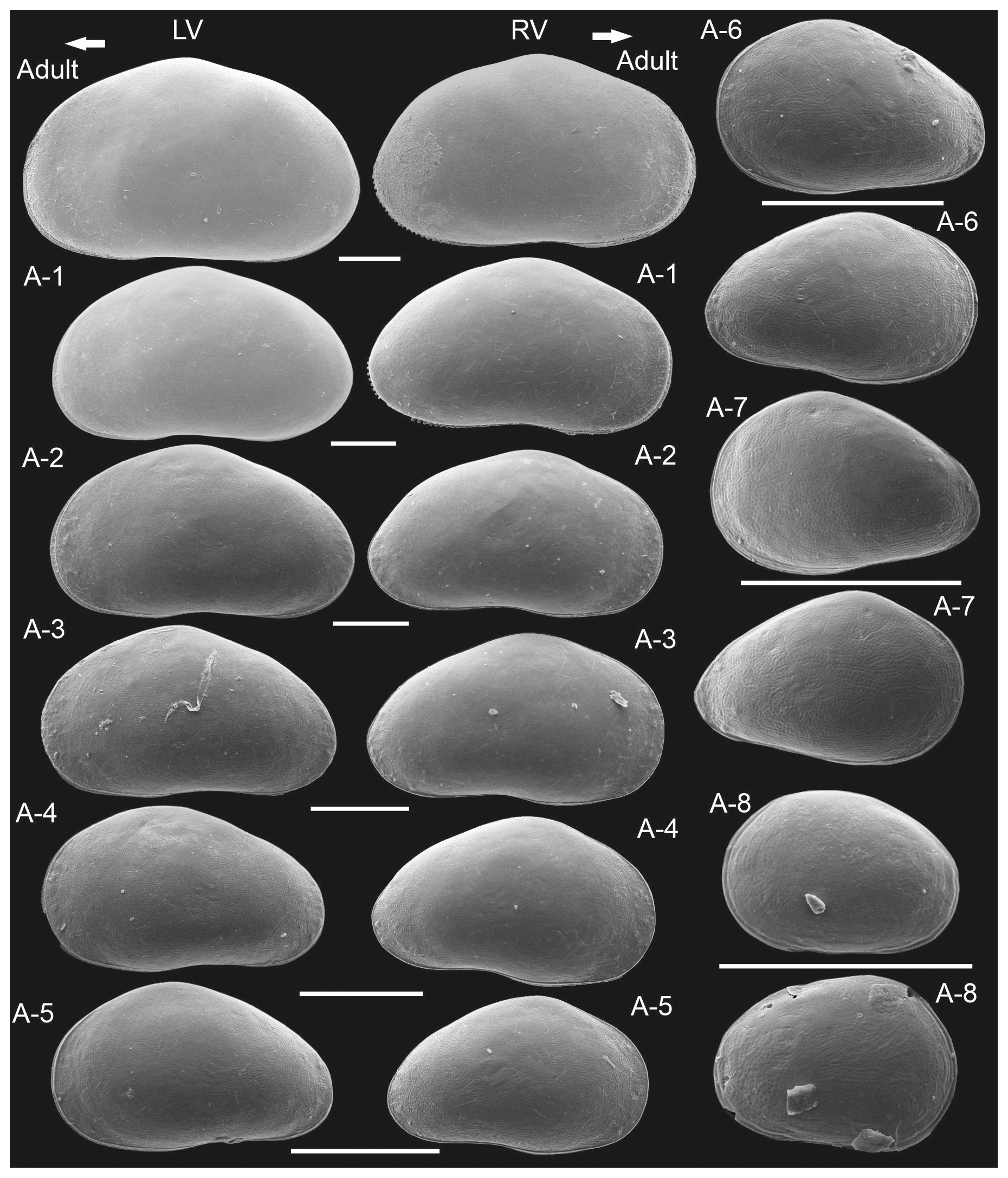
Figure 3Left (LVs) and right valves (RVs) of adult and juvenile instars (A-1 to A-8) of Heterocypris exodonta from Nam Co, Tibetan Plateau (NC-0919). All instars are shown at the largest possible magnification to highlight surface features and valve morphology. The arrow points anteriorly. Scale bars show 200 µm.
Habitat. Ephemeral pond located at the Nam Co Monitoring and Research Station for Multisphere Interactions (NAMORS). Altitude: 4728 m a.s.l. (Anslan et al., 2020).
Figure 4Close-up views of the left valves of juvenile instars of Heterocypris exodonta from Nam Co, Tibetan Plateau (NC-0919): (A) A-8, (B) A-6, (C) A-4, and (D) A-2, showing the progressive development of the reticulation pattern.
Table 3Mean values and standard deviations of length and height of right (RVs = 424) and left (LVs = 401) valves and growth ratios between successive instars of Heterocypris exodonta collected from Nam Co, Tibetan Plateau; n denotes number of valves.
-
Heterocypris incongruens (Ramdohr, 1808)
Material. Females from San Nicolás Tetelco, Mexico City, Mexico.
Reproduction. Predominantly asexual, although sexual populations, including males, have been recorded from multiple localities, particularly in Europe and North Africa (Meisch, 2000; Rossi et al., 2007), as well as in Türkiye (Yavuzatmaca and Külköylüoğlu, 2019).
Ontogeny. Egg, eight juvenile instars, and one adult stage (Fig. 5). Although a ninth juvenile instar (A-9) has been reported for this species (Rossi et al., 2015), it was not observed in the present study.
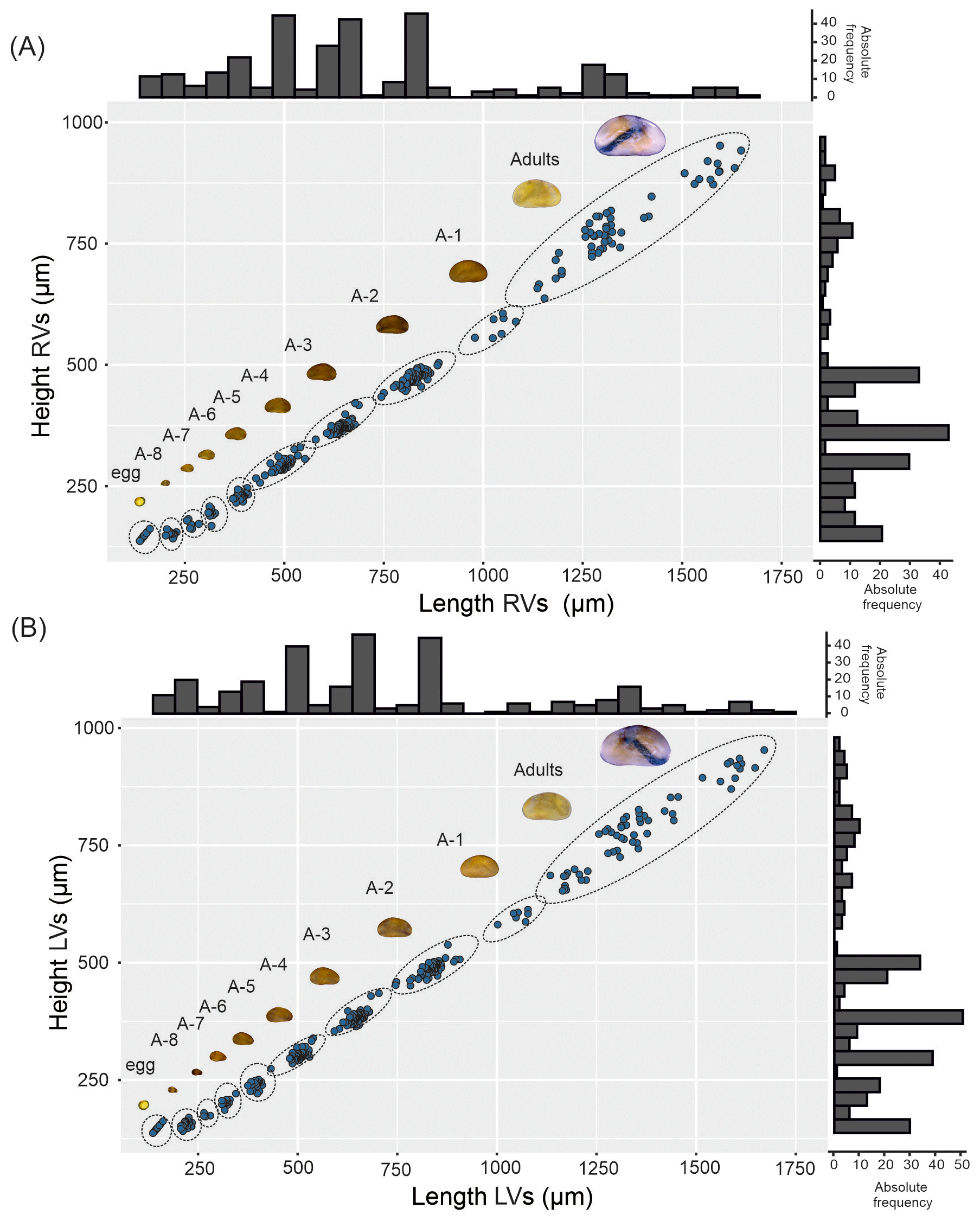
Figure 5Valve ontogeny of Heterocypris incongruens from Mexico City, Mexico (HI-M19). The scatterplot shows the length and height of (A) right valves (RVs), n=300, and (B) left valves (LVs) n=280. The histograms display the abundances of eggs, juvenile instars, and adult stage.
Egg characteristics. The eggs display an orange-colored (Fig. 6A) outer eggshell layer showing a porous appearance (Fig. 6B–D). On the surface of the egg, remnants of environmental materials, such as sediment or plant remains, are observed (Fig. 6C), providing an additional protective layer for the embryo due to adhesive properties. The inner eggshell layer envelops the embryo (Fig. 6E). Furthermore, the external surface of the eggshell shows small protuberances (Fig. 6F).
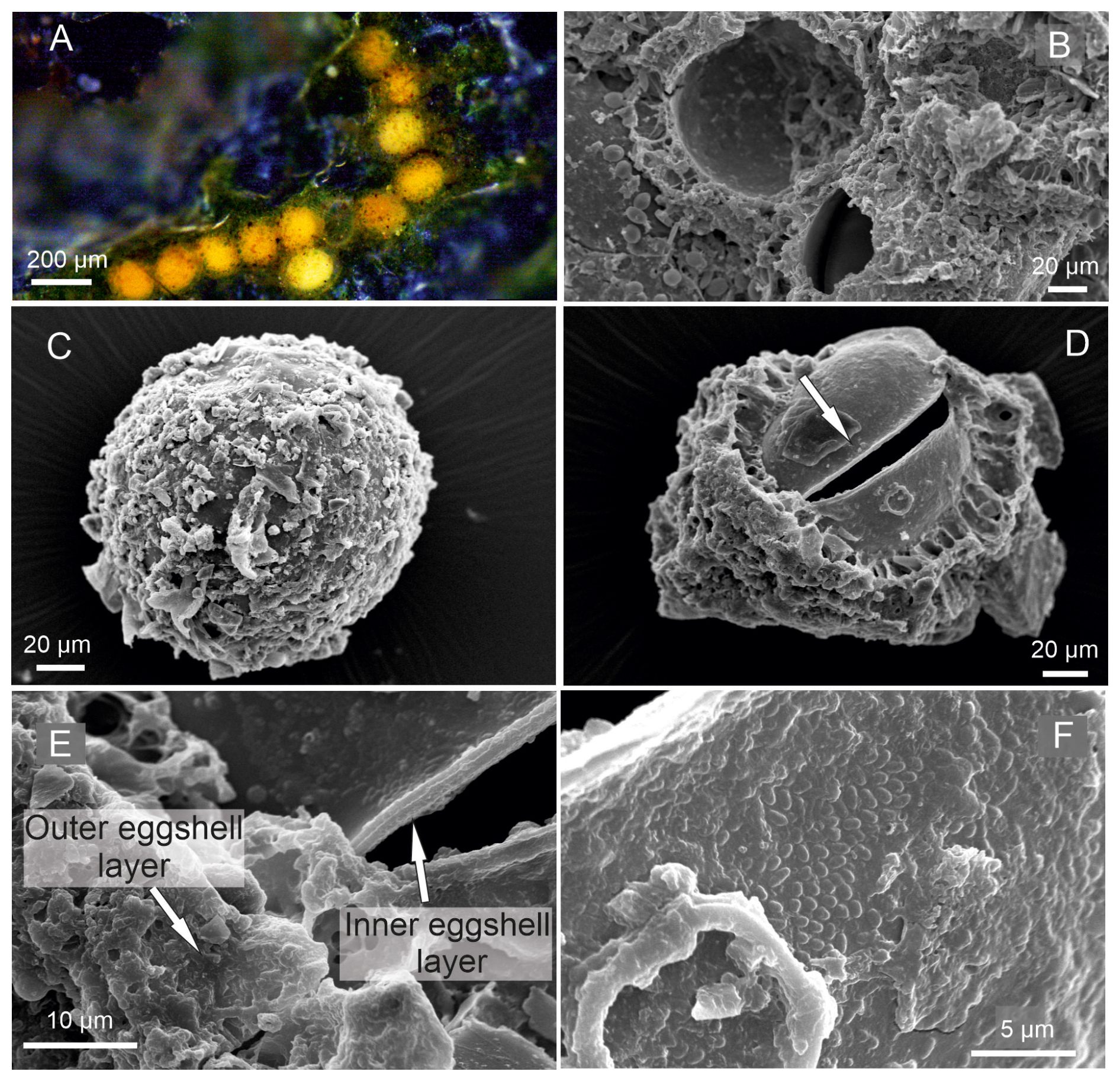
Figure 6Heterocypris incongruens. (A) Egg cluster attached to a substrate, (B) egg showing an irregular opening after hatching, (C) unhatched egg with the outer eggshell layer showing adhered external particles, (D) irregular opening indicated by the arrow, (E) close-up view of the inner and outer eggshell layers, (F) outer surface of inner layer of the eggshell displaying small protuberances.
Adult valves. The color of the valves is uniformly yellowish; however, some of the larger females were whitish in color (Fig. 5). The surface has normal pores and bumps, and the valves are elongated. The RV is smaller than the LV, and the LV overlaps the RV. The highest part is in the middle region of the valves (Fig. 7). RVs with a broad inner lamella, crenulation or tubercles in internal view on anterior and posterior edges (see Bonilla-Flores et al., 2025).
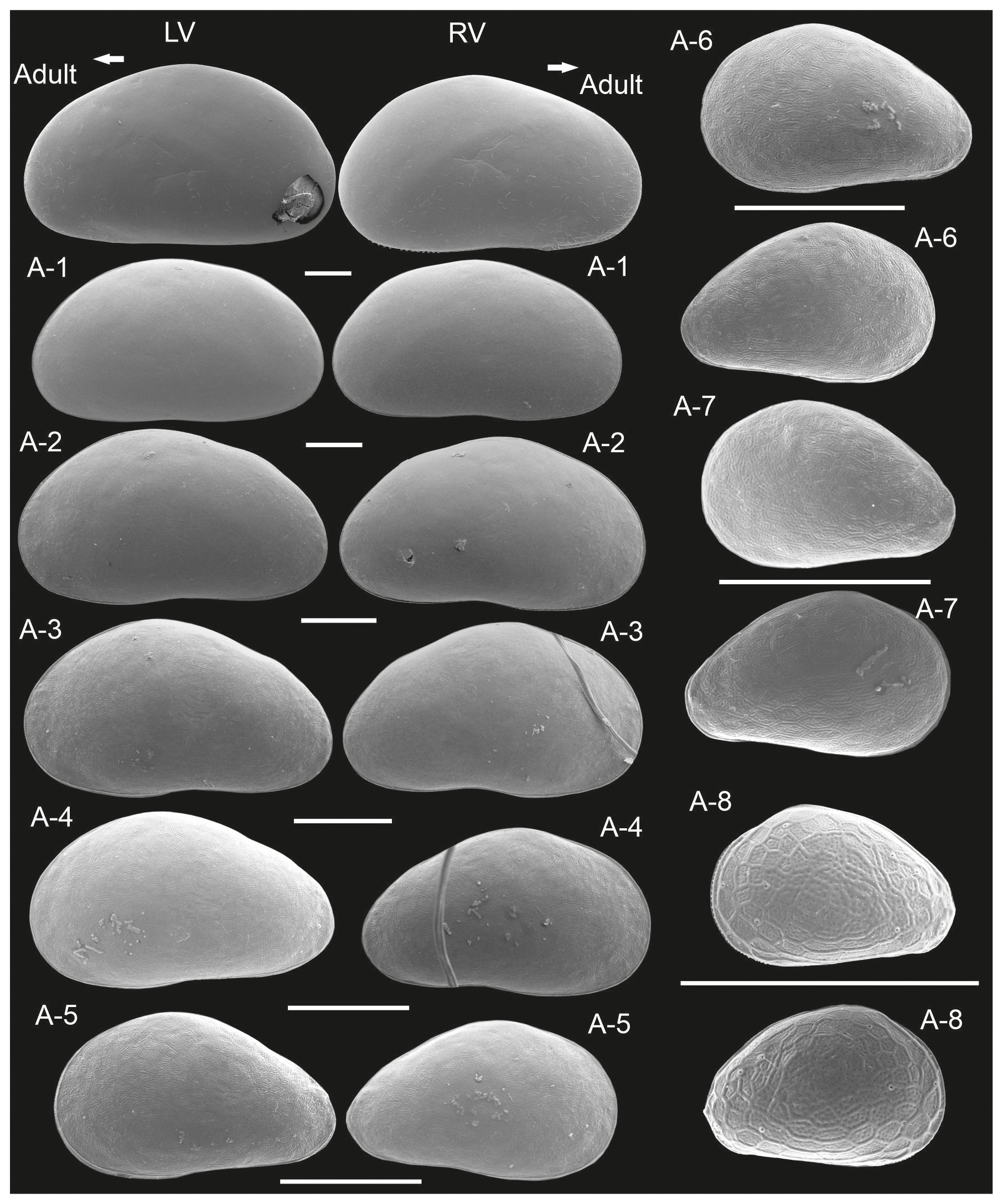
Figure 7Left (LVs) and right valves (RVs) of adult and juvenile instars (A-1 to A-8) of Heterocypris incongruens from Mexico City, Mexico (HI-M19). All instars are shown at the largest possible magnification to enhance visualization of valve morphology and surface features. The arrow points anteriorly. Scale bars show 200 µm.
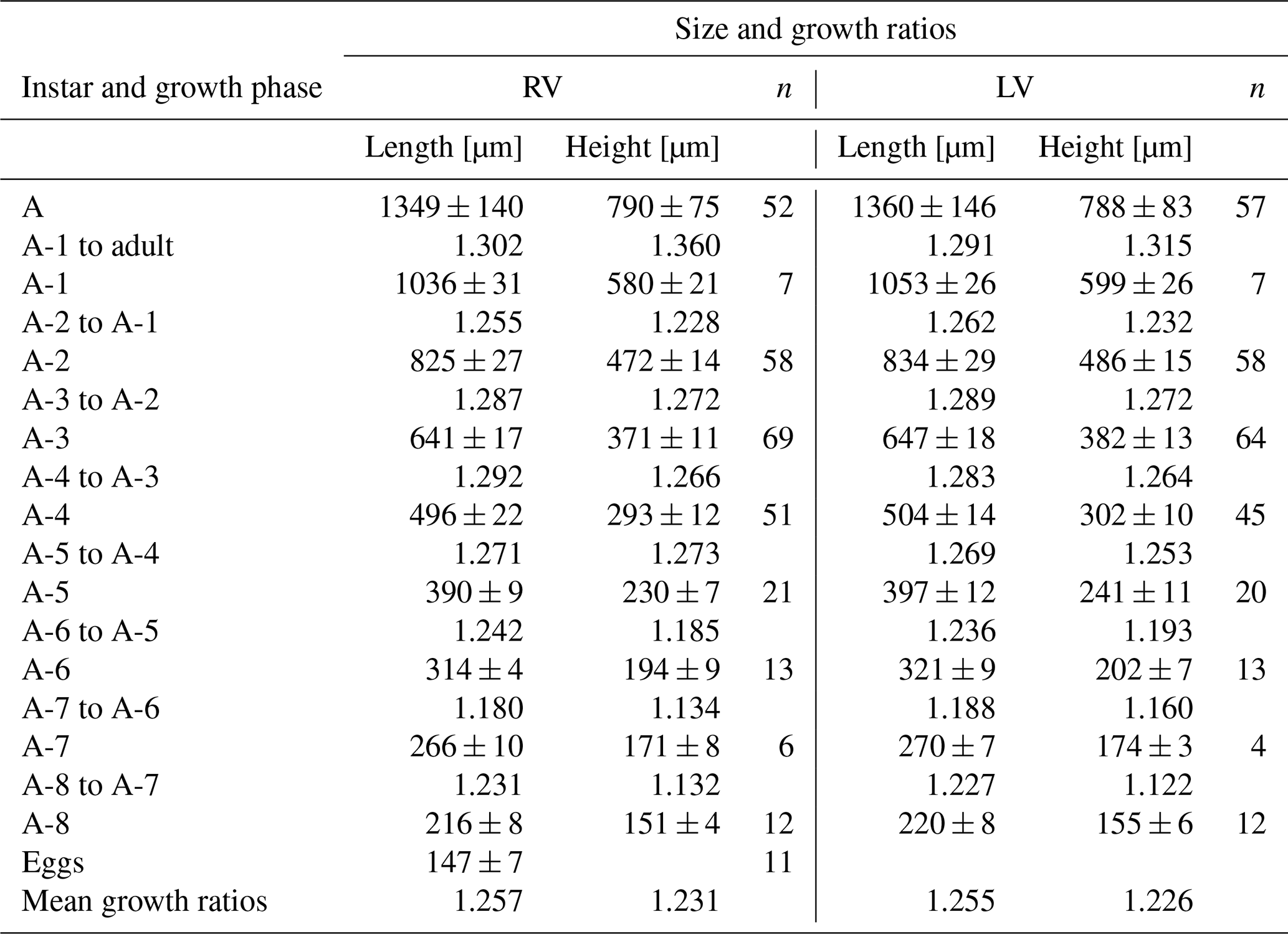
Juvenile valves (Fig. 7). Surface with normal pores with setae. The reticulation is hexagonal and includes irregular polygonal forms in A-8 and is less pronounced in the last juvenile stage (A-2) (Fig. 8). The polygonal pattern is variable, including not only hexagons but also other irregular polygonal shapes. A triangular shape was observed for stage A-8, although this form is observable in subsequent molting instars, becoming more arched dorsally in A-1. On average, A-8 juvenile RVs measure 216 µm in length and 151 µm in height, and LVs measure 220 µm in length and 155 µm in height. Adult RVs measure 1349 µm in length and 790 µm in height, and LVs measure 1360 µm in length and 788 µm in height, on average (Table 3). The mean growth ratio (length and height) for the right valves is 1.257 and 1.231, and for the left valves, it is 1.255 and 1.226, respectively (Table 3).
Habitat. Mainly temporary waterbodies (Fryer, 1997; Meisch, 2000; Vandekerkhove et al., 2012).
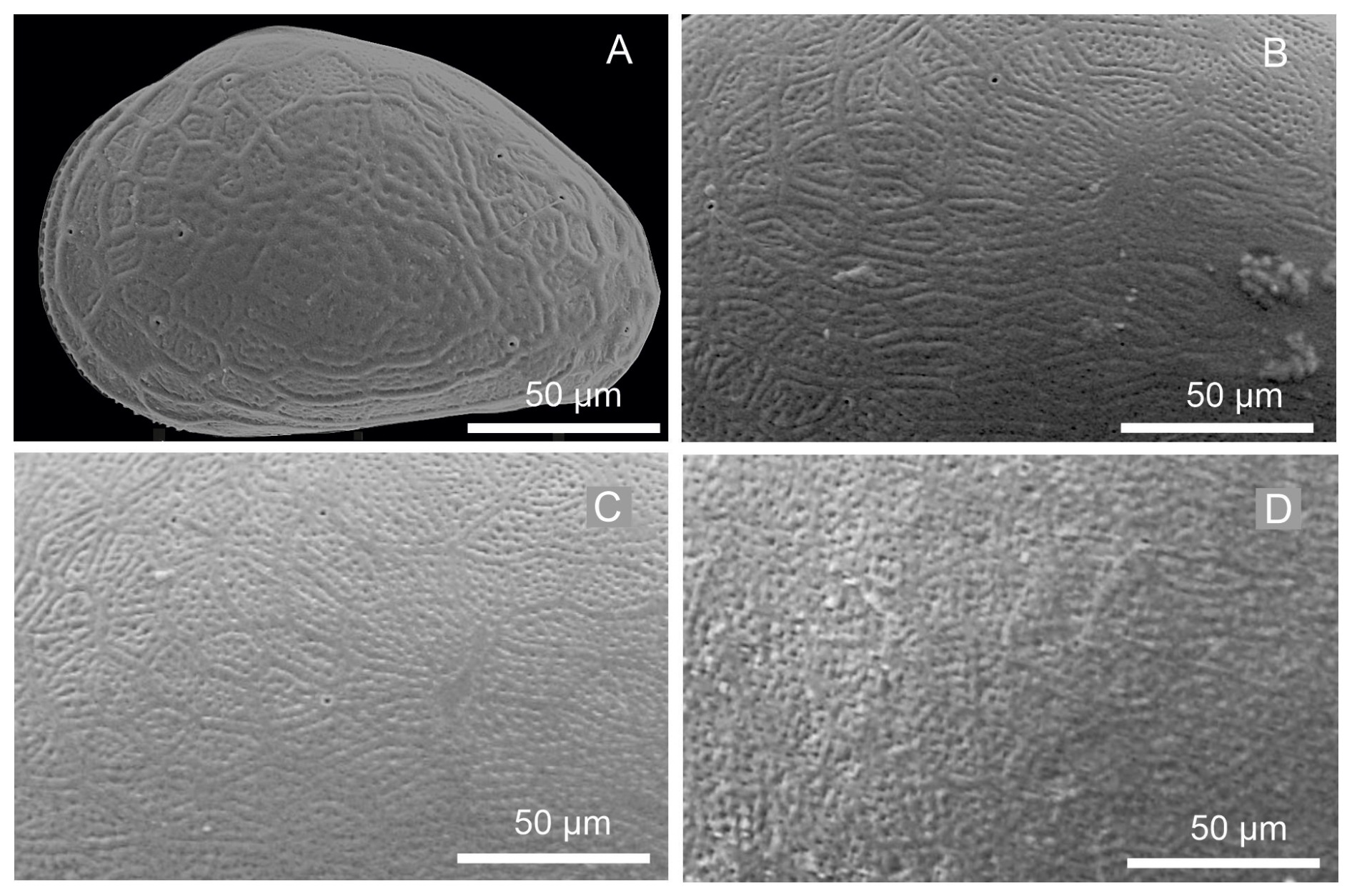
Figure 8Close-up views of the left valves of juvenile instars: (A) A-8, (B) A-6, (C) A-4, and (D) A-2, showing densely reticulated surface pattern.
Table 4Mean values and standard deviations of length and height of right (RVs = 300) and left (LVs = 280) valves and growth ratios between successive instars of Heterocypris incongruens collected from Mexico City; n denotes number of valves.
-
Heterocypris salina (Brady, 1868)
Material. Females were collected from the Botanical Garden, Braunschweig, Germany.
Reproduction. Asexual populations from Braunschweig, Germany. Males and females were recorded from Crete, Greece (Petkowski et al., 2000).
Ontogeny. Egg, eight juvenile instars, and one adult stage (Fig. 9).
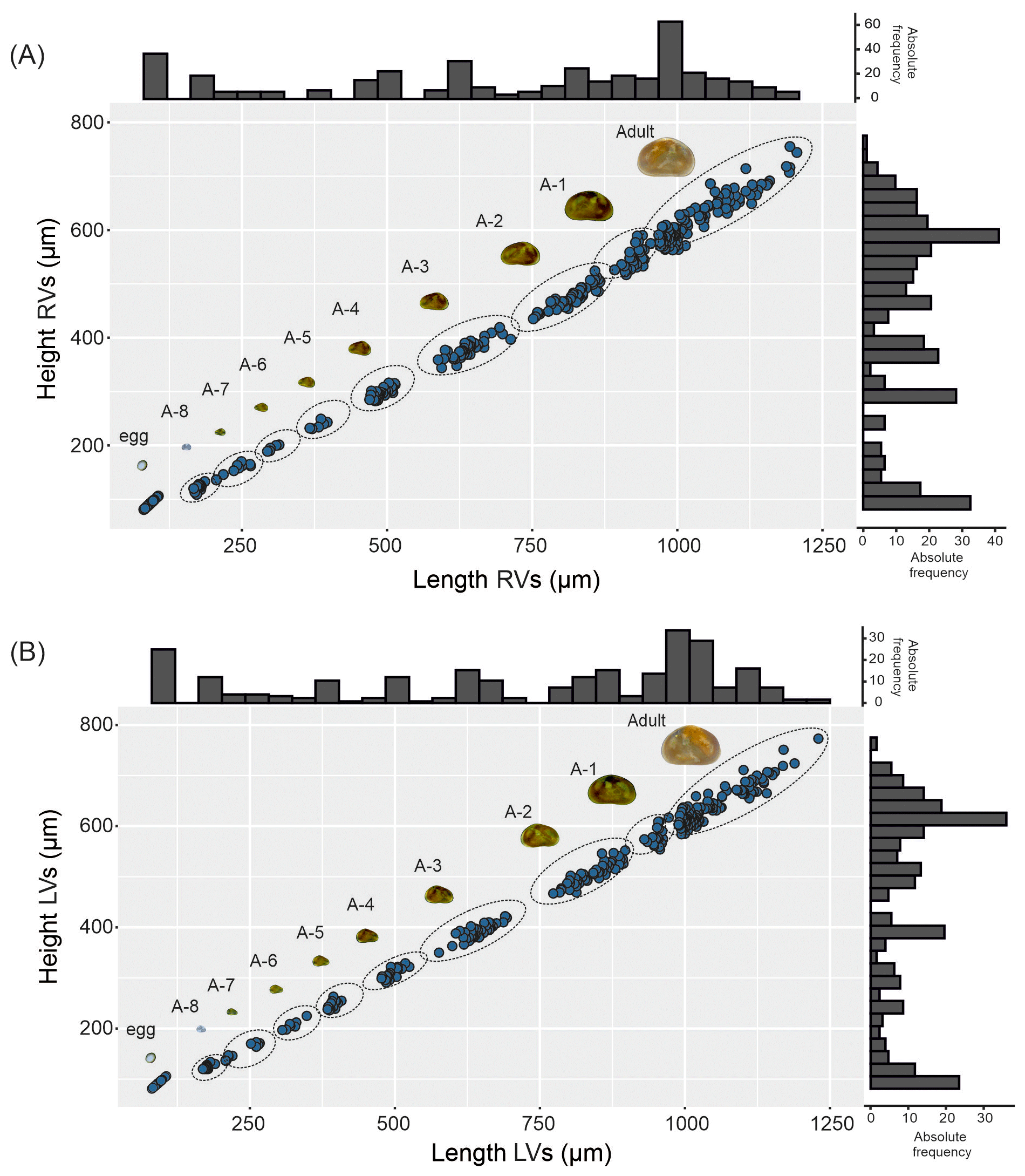
Figure 9Valve ontogeny of Heterocypris salina from Braunschweig, Germany (HS-G19). The scatterplot shows the length and height of (A) right valves (RVs), n=300, and (B) left valves (LVs), n=287. The histograms display the abundances of eggs, juvenile instars, and the adult stage.
Egg characteristics. Eggs exhibit two colorations, either yellowish or whitish (Fig. 10A). They are laid in clusters, adhering to each other through adhesive secretions produced during oviposition (Fig. 10B–D). The eggs are surrounded by an inner eggshell layer and an outer eggshell layer (Fig. 10E–F).
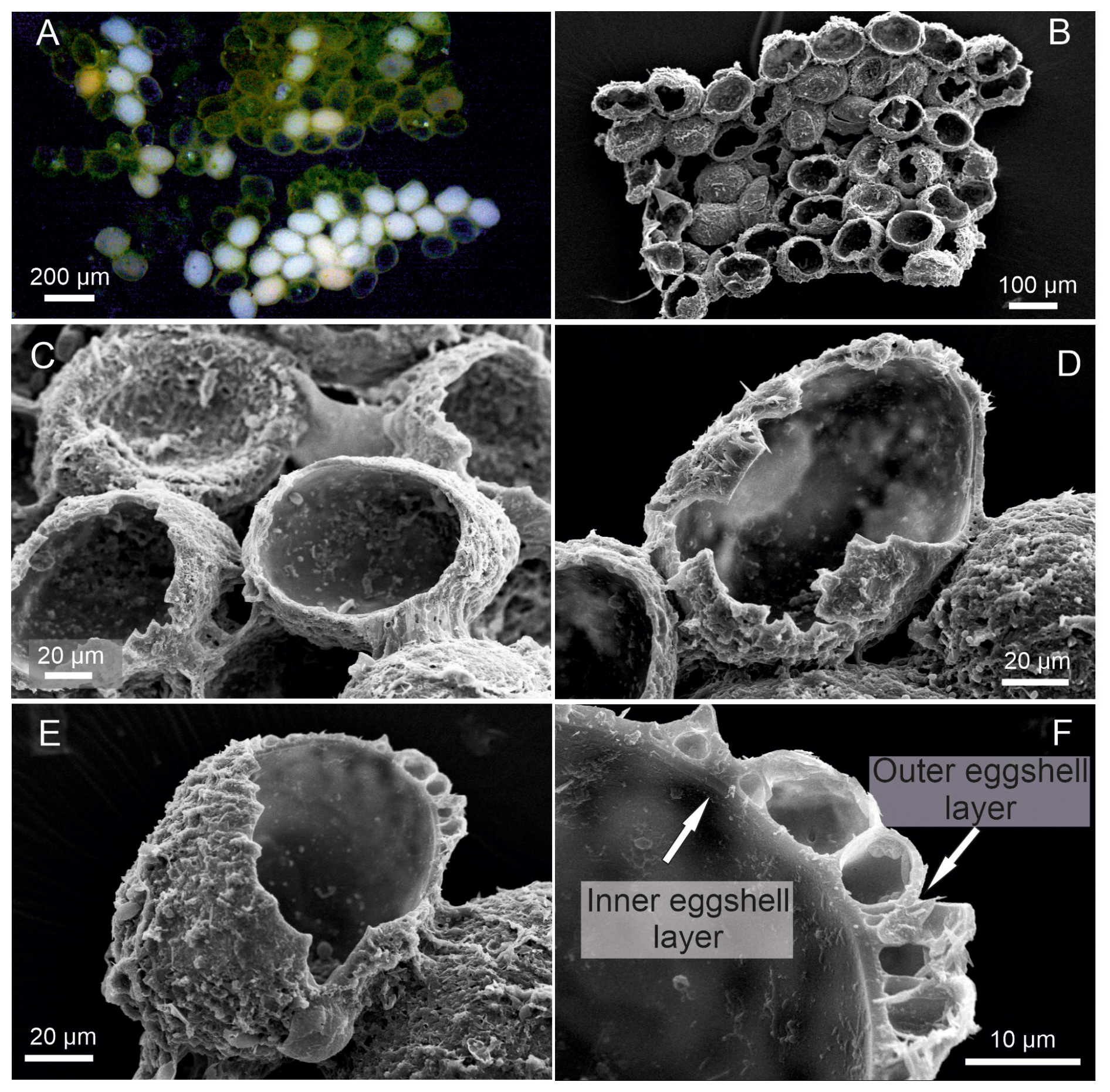
Figure 10Heterocypris salina. (A–B) Egg clusters, (C) adhesive material between eggs, (D–E) eggs showing an irregular opening after hatching with exposed eggshell layers, (F) close-up view of the eggshell illustrating the inner and outer eggshell layers.
Adult valves. Both valves display a brown coloration pattern, with a pair of lighter perpendicular lines (Fig. 9). The external surface has normal pores with a seta. Compressed and triangular in the dorsal region (Fig. 11). The LV is longer than the RV, overlapping the RV. The external surface of the RV has inconspicuous crenulation along the anterior and posterior margins; detailed high-magnification images of this feature are provided in Bonilla-Flores et al. (2025).

Figure 11Left (LVs) and right valves (RVs) of adult and juvenile instars (A-1 to A-8) of Heterocypris salina from Braunschweig, Germany (HS-G19). All instars are shown at the largest possible magnification to enhance visualization of valve morphology and surface features. The arrow points anteriorly. Scale bars show 200 µm.
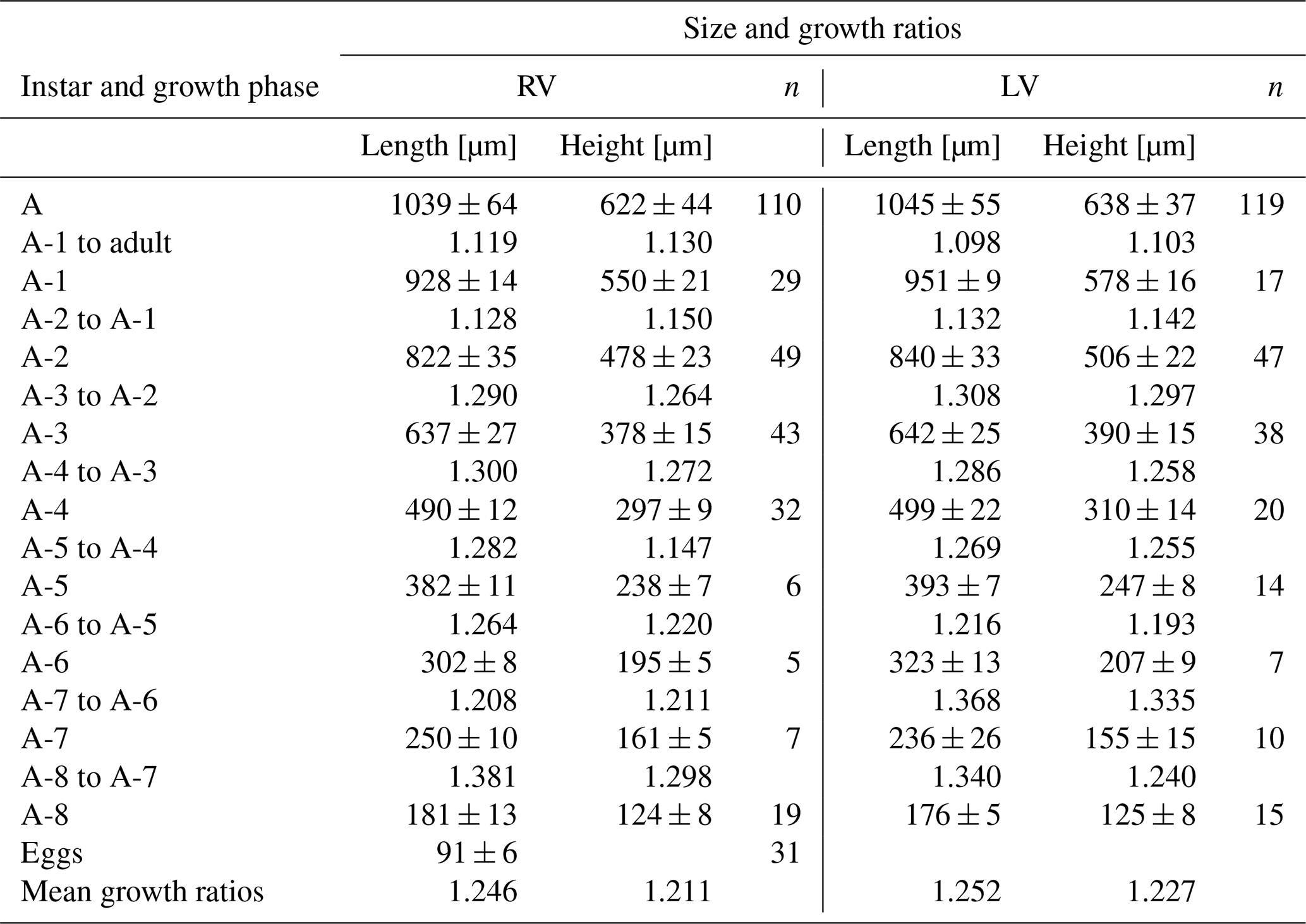
Juvenile valve. External surface with normal pores and setae. The irregular reticulation in surface valves gradually dissipates in the last juveniles (Fig. 12). The shape of stage A-8 is rounded, which then transitions into a more triangular form in the subsequent molting instars to become more arched in A-1; in this stage, small tubercles are also observed along the postero-dorsal margin of the RV (Fig. 11). On average, A-8 juvenile RVs were 181 µm long, and their height was 124 µm, while the LVs were 176 µm long and 125 µm high. Adult RVs were 1039 µm long and 622 µm high; LVs were 1045 µm long and 638 µm high (Table 4). The mean growth ratio (length height) for RVs was 1.246 and 1.211, and for LVs, it was 1.252 and 1.227, respectively (Table 4).
Habitat. Eurytopic species; tolerates high values of water conductivity and organic pollution; found in shallow ponds and temporary pools (Mezquita et al., 1999; Meisch, 2000).
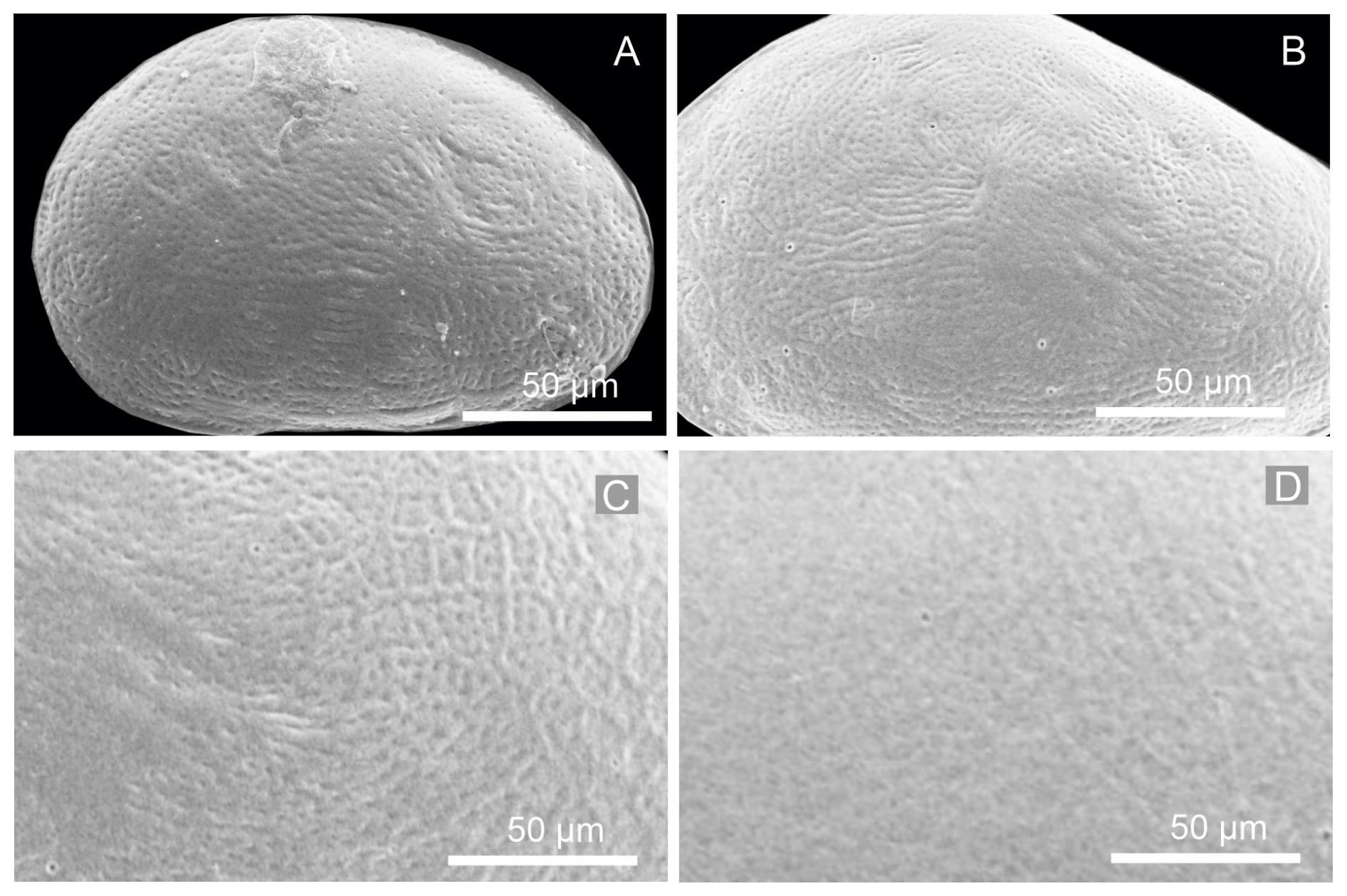
Figure 12Close-up views of the left valves of juvenile instars: (A) A-8, (B) A-6, (C) A-4, and (D) A-2, showing the reticulated surface pattern, which becomes progressively less pronounced and more diffuse in the A-2 instar.
Table 5Mean values and standard deviations of length and height of right (RVs = 331) and left (LVs = 287) valves and growth ratios between successive instars of Heterocypris salina collected from Braunschweig, Germany; n denotes number of valves.
5.1 Egg morphology and ecological significance
It was observed that the eggs of the three Heterocypris species share a remarkably similar overall morphology, including an ellipsoidal shape, comparable size, and a relatively resistant external appearance. These similarities suggest a shared adaptation to ephemeral aquatic environments. However, surface features are not uniform across ostracod taxa; for instance, Chlamydotheca arcuata (Sars, 1901) exhibits tuberculate ornamentation (Díaz and Lopretto, 2017), indicating that broader comparative studies are required to better understand the diversity and evolutionary significance of egg morphology.
The eggs of Heterocypris exodonta, H. incongruens, and H. salina have two eggshell layers, with the outer layer displaying a rigid-looking roughness, which is vital for protecting the embryo. The structure of ostracod eggs has been described in classical histological studies, which demonstrate that the eggshell is formed during oogenesis and consists of two layers, including an inner layer produced by the oocyte and an outer layer secreted by the ovarian epithelium (Woltereck, 1898; Weygoldt, 1960; Hartmann, 1968). In this context, the interpretation of eggshell morphology based solely on SEM observations should be approached with caution. Features such as a rough or porous surface may not represent primary structural characteristics but instead result from post-depositional processes, including water uptake and expansion of the outer layer (Wohlgemuth, 1914).
Furthermore, early embryological studies have demonstrated that ostracod structures arise through progressive differentiation and proliferation of tissues rather than as static or discrete layers (Müller-Calé, 1913). This dynamic developmental process may lead to misleading structural interpretations when observations are based solely on surface imaging techniques.
Consequently, previously described “layers” based on SEM observations (e.g., Dumont et al., 2002; Özuluğ and Suludere, 2012) for Heterocypris incongruens should be reconsidered as they may reflect optical or structural artifacts rather than true biological organization. Similarly, particles or external coatings observed on the egg surface are not intrinsic components of the eggshell but are likely associated with adhesive secretions produced during oviposition. These findings highlight the importance of integrating classical histological knowledge with modern imaging techniques to avoid misinterpretations of ostracod egg morphology. Despite these limitations, the eggs of Heterocypris species are known to exhibit high resistance to environmental stressors, including desiccation and extreme temperatures (−18 to 42 °C), and can remain viable in a dormant state for over 20 years (Angell and Hancock, 1989). Future experimental studies should focus on testing egg resistance under controlled environmental conditions, including temperature extremes and simulated drought, to better understand hatching success, survival, and developmental rates.
In ecological terms, resistant eggs play a key role in the persistence of ostracod populations in temporary and unpredictable environments. However, their production and hatching dynamics are not controlled by temperature alone. Experimental studies on Heterocypris incongruens have shown that both temperature and photoperiod influence the proportion of subitaneous versus diapausing eggs, as well as their hatching timing and success (Rossi et al., 1996, 2013). In addition, temperature affects growth, development, and fecundity in complex and often non-linear ways rather than acting as a single deterministic factor (Aguilar-Alberola and Mesquita-Joanes, 2014). These environmental cues interact with maternal effects and genotype-specific responses, influencing egg size, structure, and hatching phenology and contributing to variability in life history strategies (Rossi et al., 2013).
In this study, eggs of H. exodonta, H. incongruens, and H. salina exhibited clear interspecific differences in size, with mean values of 129 ± 7 µm (n=20), 147 ± 7 µm (n=11), and 91 ± 6 µm (n=31), respectively. These differences are noteworthy given that all species were maintained under the same laboratory conditions (18–23 °C; 14:10 h light: dark photoperiod), suggesting that egg size variation reflects intrinsic species-specific traits rather than environmental variation alone. However, previous studies have demonstrated that egg size and hatching phenology in Heterocypris can be modulated by environmental conditions, particularly photoperiod, as well as maternal effects (Rossi et al., 2013). Therefore, although the controlled conditions in this study allow for direct comparison among species, they may not capture the full extent of phenotypic plasticity observed in natural populations.
Furthermore, resting eggs accumulate in sediments, forming egg banks, which act as ecological reservoirs that enhance resilience and facilitate recolonization after disturbance (Rosa et al., 2020). These egg banks contribute to metapopulation dynamics by enabling local extinction and recolonization processes (Rossi and Menozzi, 2012) and may be spatially redistributed by hydrological processes such as flooding (Rosa et al., 2020). In addition, hatching patterns are influenced by species traits and phylogenetic relationships, as well as environmental cues, resulting in staggered emergence and promoting population persistence under fluctuating conditions (Rosa et al., 2021). The persistence of dormant stages, together with broader “ostracod banks”, further enhances survival and recolonization capacity in highly variable habitats (Wang et al., 2025).
Finally, it is important to consider that the egg characteristics observed in this study may be influenced by laboratory conditions, including constant temperature and photoperiod regimes. Although these conditions were necessary to ensure comparability among species and to meet the objectives of describing valve ontogeny, characterizing egg morphology and evaluating growth ratios, they may affect egg production, size, structure, and diapause expression, as shown in previous experimental studies (Rossi et al., 1996, 2013). Therefore, the results should be interpreted with caution when extrapolating to natural populations. Despite this limitation, our study provides a comparative framework for understanding interspecific differences in egg morphology and developmental patterns among closely related species from distinct geographic regions. Further studies are needed to evaluate how environmental variability influences egg traits, viability, and hatching success, as well as the physiological and genetic mechanisms underlying these responses (Bellin et al., 2020; Horne and Martens, 1998; Delorme, 2011).
5.2 Ontogeny and taxonomic implications
The ontogenies of H. exodonta, H. incongruens, and H. salina (Fig. 13), in terms of the number of molting instars of the valves, are consistent with the general pattern described for podocopid ostracods. All three species exhibit eight juvenile instars followed by a single adult stage, in agreement with previous observations from Marburg, Germany (Schreiber, 1922); Bogotá, Colombia (Roessler, 1983); Büyükçekmece Lake, Istanbul (Kubanç et al., 2007); and temporary pools on the Iberian Peninsula (Aguilar-Alberola and Mezquita, 2008). However, deviations from this standard nine-stage ontogenetic pattern have been reported in some Heterocypris species. In particular, the presence of an additional early developmental stage (A-9), interpreted as a prenaupliar stage, has been documented in H. bogotensis, H. bosniaca, and H. incongruens (Roessler, 1983; Aguilar-Alberola and Mesquita-Joanes, 2013; Rossi et al., 2015), indicating that ontogenetic patterns within the genus may be more variable than traditionally assumed. In our study, this additional stage was not observed, which may be related to sampling limitations or to the difficulty of detecting early developmental stages, given the fragility of the exuviation process and the rapid succession of molts.
Ontogenetic analysis remains a fundamental tool in ostracod taxonomy as morphological changes through successive instars provide key characters for species identification and for interpreting phylogenetic relationships (Brooks, 1886; Shaver, 1953; Watabe and Kaesler, 2004; Boomer et al., 2003; Danielopol et al., 2008). In this context, our study extends previous work by providing a complete ontogenetic framework for H. incongruens and H. salina based on SEM observations. Although Schreiber (1922) presented detailed drawings of H. incongruens, high-resolution documentation of surface features across all instars has remained limited until now.
Our SEM images show that valve ornamentation changes throughout development rather than remaining static. Reticulation patterns have also been reported in other non-marine ostracods, including Cypris pubera (Yousef et al., 2024), Chlamydotheca arcuata (Díaz and Lopretto, 2017), Eucypris virens (Smith and Martens, 2000), and Strandesia bicuspis (Liberto et al., 2014), and in some candonids such as Candona xizangensis (Akita et al., 2016) and Fabaeformiscandona myllaina (Smith and Kamiya, 2007). In the three Heterocypris species studied here, reticulation follows a clear ontogenetic trend: early instars (A-8 to A-2) display a well-developed and variable polygonal pattern, including hexagonal and irregular forms, which becomes progressively weaker in later stages. This pattern is consistent across H. exodonta, H. incongruens, and H. salina, with minor interspecific differences in terms of its persistence, and is comparable to that observed in juvenile instars of H. bosniaca (Aguilar-Alberola and Mezquita, 2008; Aguilar-Alberola and Mesquita-Joanes, 2013). Taken together, these results indicate that valve ornamentation alone is not a reliable taxonomic character, but it becomes informative when evaluated in relation to developmental stage.
Despite being widely distributed and eurytopic species (Meisch, 2000), detailed ontogenetic descriptions for H. incongruens and H. salina have been scarce, particularly regarding fine surface morphology. Our results, therefore, contribute new data that refine the morphological characterization of these species and improve the identification of juvenile stages, which are often problematic in taxonomic studies.
Overall, our findings highlight the importance of integrating ontogenetic analysis with high-resolution imaging techniques to better understand morphological variability and improve species discrimination. However, given the observed variability in developmental patterns and valve ornamentation, ontogenetic characters should be used cautiously and in combination with other morphological or molecular evidence.
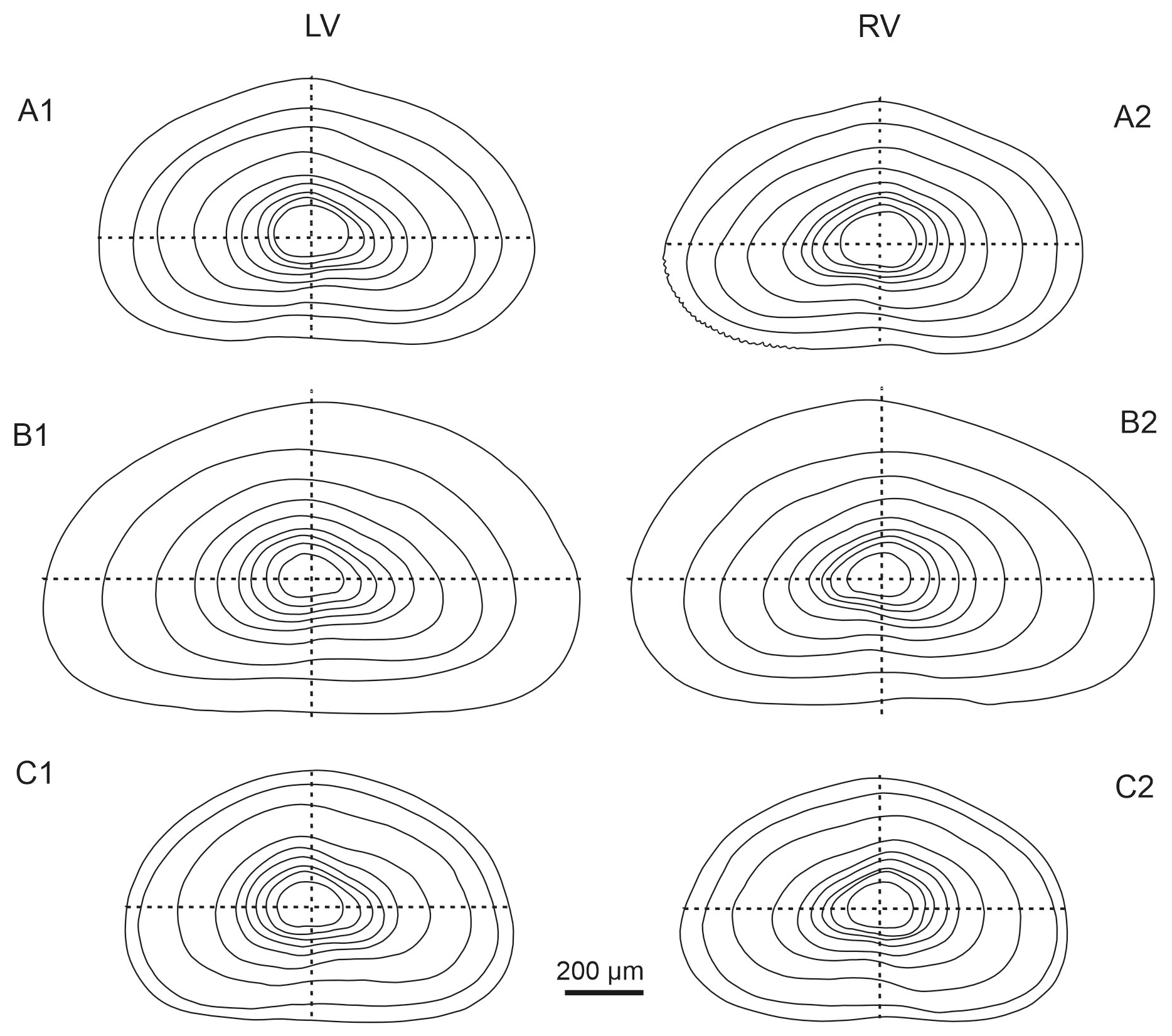
Figure 13Comparison of external valve outlines across nine ontogenetic instars (from juvenile A-8 to adult). Left (LVs) and right (RVs) valves of (A1–A2) Heterocypris exodonta, (B1–B2) H. incongruens, and (C1–C2) H. salina.
The frequent occurrence of H. incongruens and H. salina (Meisch, 2000) has long been recognized. However, despite their prevalence, detailed comparative analyses of these species are missing. The contour diagrams presented herein represent a significant step forward in elucidating the ontogenetic development of these ostracod species, providing valuable insights into their morphological similarities and differences. By comparing the outlines of the valves (Fig. 13), differences in shapes among species become apparent, particularly in early instars. In A-8, H. exodonta and H. salina display a more rounded outline, whereas H. incongruens tends to exhibit a more triangular shape. Although early instars may be more flexible and potentially prone to deformation, no clear evidence of preparation artifacts was observed in our material. Therefore, the observed differences are more likely to reflect interspecific variation rather than deformation during SEM processing. However, this interpretation should still be approached with caution as previous observations by Schreiber (1922) show a more rounded outline for the right valve of H. incongruens, suggesting that some degree of variability may occur. In addition, the presence of cryptic species within H. incongruens may contribute to subtle differences in early ontogenetic stages. Apart from this consideration, the outlines of later instars are more consistent and closely resemble the triangular or conical shapes observed in this study. Furthermore, the A-5 to A-3 juvenile valves of H. salina feature a notably rounded dorsal region, unlike the straighter profiles of H. exodonta and H. incongruens. The contour diagrams presented here will serve as crucial benchmarks for future studies, providing a comprehensive understanding of the ontogenetic development of these ostracods and facilitating comparisons with related species.
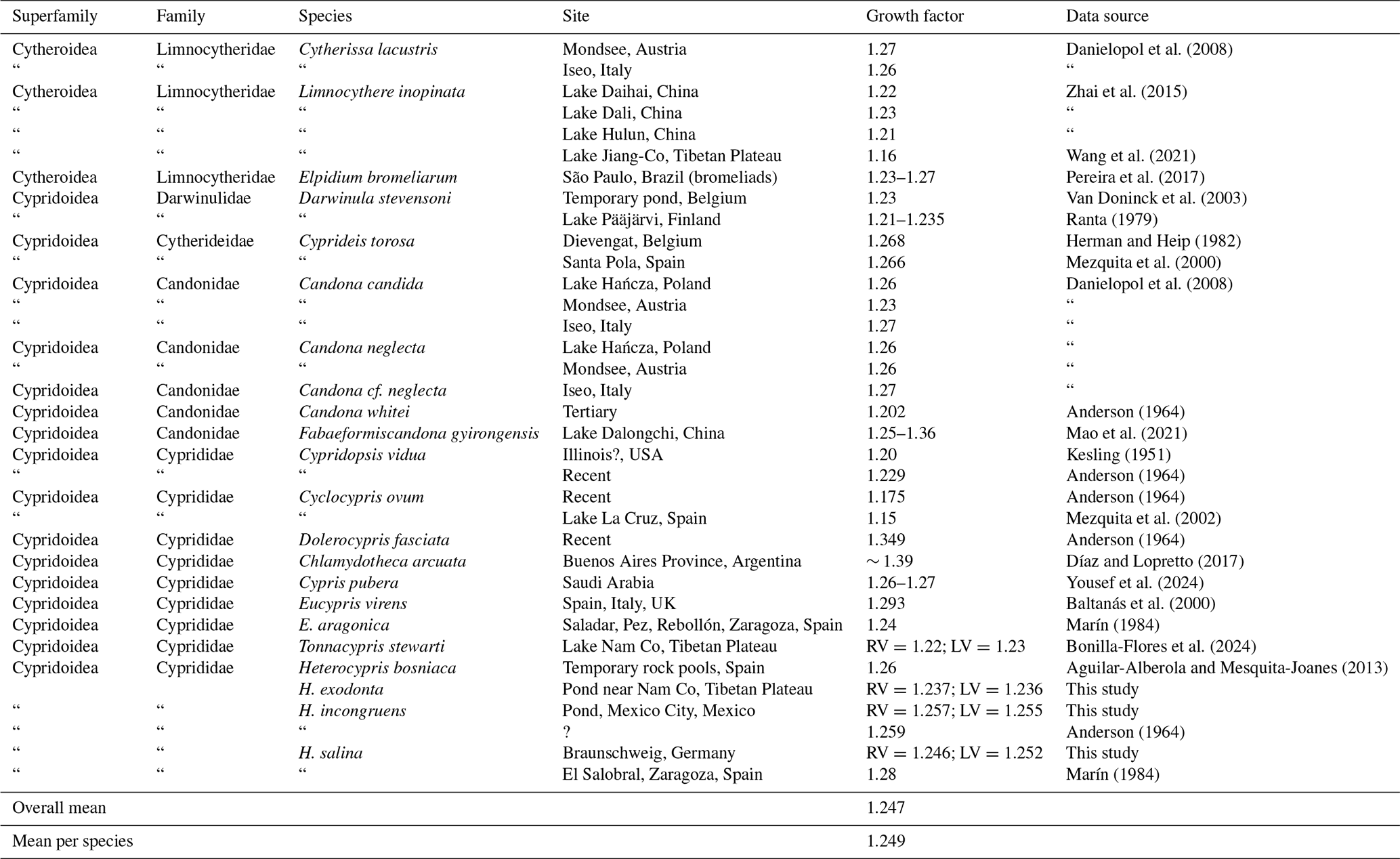
Our dataset shows that growth ratios in the three species studied converge on average values close to those predicted by Brooks' rule for crustaceans (∼ 1.26). However, rather than indicating a strictly predictable pattern, the data reveal considerable variability both among species and across developmental stages. This variability is also evident in the literature, where growth factors span a relatively wide range (Table 6), suggesting that geometric growth represents a general tendency rather than a fixed rule. Although subtle, the observed differences in growth ratios among the studied species do not provide sufficiently consistent information to serve as a reliable diagnostic character for distinguishing closely related taxa with minimal morphological differences. Therefore, confirming the taxonomic utility of growth ratios remains challenging and requires additional data from closely related species, as well as broader geographic and environmental sampling (Brooks, 1886; Watabe and Kaesler, 2004). Furthermore, as noted by Kesling (1952), growth factors should be interpreted with caution as increases in valve size may lead to variations in growth values, and adult stages may differ functionally and morphologically from juvenile instars. In particular, the larger increment observed during the final molt has been attributed to the development of reproductive structures at maturity, which may contribute to deviations from earlier ontogenetic growth patterns (Kesling and Crafts, 1962).
Table 6Growth factors for non-marine ostracod species from this study are compared with data from the literature. Quotation marks (“) indicate repetition of the species name and reference. The symbol (?) indicates that information is uncertain or not available.
The life cycle study of Darwinula stevensoni living in a temporary pond in Belgium from spring to summer revealed a valve length growth ratio of 1.236. The authors suggest that temperature was identified as an important factor influencing developmental timing as higher temperatures were associated with faster development and earlier reproductive maturity (Van Doninck et al., 2003). Similar effects of temperature on growth, including size and growth ratios, have been documented in Heterocypris, where thermal conditions influence developmental rates and life history traits (Aguilar-Alberola and Mesquita-Joanes, 2014). In our study, the three Heterocypris species were cultured at 18–23 °C and exhibited relatively short life cycles (approximately 35–60 d). However, these observations should be interpreted with caution as temperature interacts with other factors such as photoperiod and intrinsic biological variability, which may also influence growth and development. Likewise, in personal observations, life cycles are slower during winter, with few molts from juveniles to adults in this season. It appears to be the case that juveniles (A-3 to A-1) enter a dormant phase in which they do not molt until they receive an environmental stimulus that signals for them to molt again; in this case, it could be the temperature acquired from sunlight. The observation was made only when the ostracods were near the window where the sunlight was strongest.
Ostracods are opportunistic feeders, and food quality can influence growth and molting (Schmit et al., 2007). In our laboratory observations, increased bacterial growth under winter light conditions may indicate stable culture conditions and frequent molting, suggesting that microbial availability may support these processes, although this relationship requires further investigation. Additionally, it is important to test for competition and potential cannibalism within the population under food-limited conditions as individuals may resort to cannibalism when alternative food sources are scarce (Herman and Heip, 1982).
For species with wide distributions, ontogenetic processes should be described and documented more in the future to fully understand morphological variations in valves and the role of different environmental factors (e.g., ionic concentration and composition, temperature, and dissolved oxygen) (Song et al., 2023) and their effects on development.
Shaver (1953) suggested using growth factors for the taxonomic characterization of supra-specific groups, underscoring the indispensability of ontogenetic studies for resolving questions related to ancestry between taxa. By identifying the similarities and differences in ontogenetic trajectories of ostracod species, these studies can provide insights into their evolutionary connections (Smith and Kamiya, 2002). However, the number of species for which these growth factors and intra-specific variability are known is still limited.
Limited variation in growth patterns may reflect shared evolutionary relationships; however, such patterns are also influenced by factors such as adult body size and egg size, which can vary across ostracod groups. As suggested by the comparative data in Table 6, differences among major taxa (e.g., Lymnocytheridae, Darwinulidae, Candonidae, and Cyprididae) may therefore be better understood in the context of both phylogenetic relationships and size-related constraints.
However, it is important to note that the degree of variation alone may not always indicate genetic connections as environmental factors can also influence growth patterns (Oakley et al., 2012; Vences et al., 2024). Finally, exploring ontogeny unveils how specific morphological traits have evolved. This information is essential for elucidating the evolutionary history of these microcrustaceans, contributing to a comprehensive assessment of their significance in aquatic ecosystems and biodiversity (Horne et al., 2004).
Our contributions include SEM photographs detailing valve surface morphology and ontogenetic changes in three Heterocypris species. Beyond their taxonomic value, our results provide insight into growth patterns in ostracods. As organisms with determinate growth, ostracods reach a fixed adult size after the final moult, and growth ratios are influenced by multiple factors, including the relationship between egg size and final body size. Comparative data (Table 6) show that growth factors vary across taxa, with larger species generally tending to exhibit higher values. For example, high growth factors occur in large-bodied species such as Eucypris virens (1.293) and Dolerocypris fasciata (1.349), whereas smaller taxa such as Cyclocypris ovum show lower values (1.15–1.175). In our dataset, H. incongruens, the largest species studied, also shows the highest mean growth ratio (RV = 1.257; LV = 1.255). A comparison between egg size and adult size (Fig. 14) shows a general positive tendency but not a strict proportional relationship, indicating that additional factors beyond initial size influence growth. In ostracods, available evidence suggests that reproductive traits such as egg number or egg volume may be related to female size, although these patterns are not always consistent. For example, a positive relationship between egg number and carapace length has been reported, whereas egg volume may be independent of female size in some species (Yousef and Alahmadi, 2025). Therefore, the relationship between egg size and maternal size remains insufficiently resolved in ostracods, particularly across species, and should be interpreted with caution. Growth ratios are also not constant throughout ontogeny. In our data, higher values occur in intermediate instars, followed by lower values in the final molts. Similar patterns have been reported previously (Liberto et al., 2014; Aguilar-Alberola and Mezquita, 2008), indicating that growth in ostracods is stage-dependent rather than uniform. Overall, growth ratios reflect a combination of developmental constraints, body size scaling, and ontogenetic variation and should not be considered to be fixed species-specific constants.
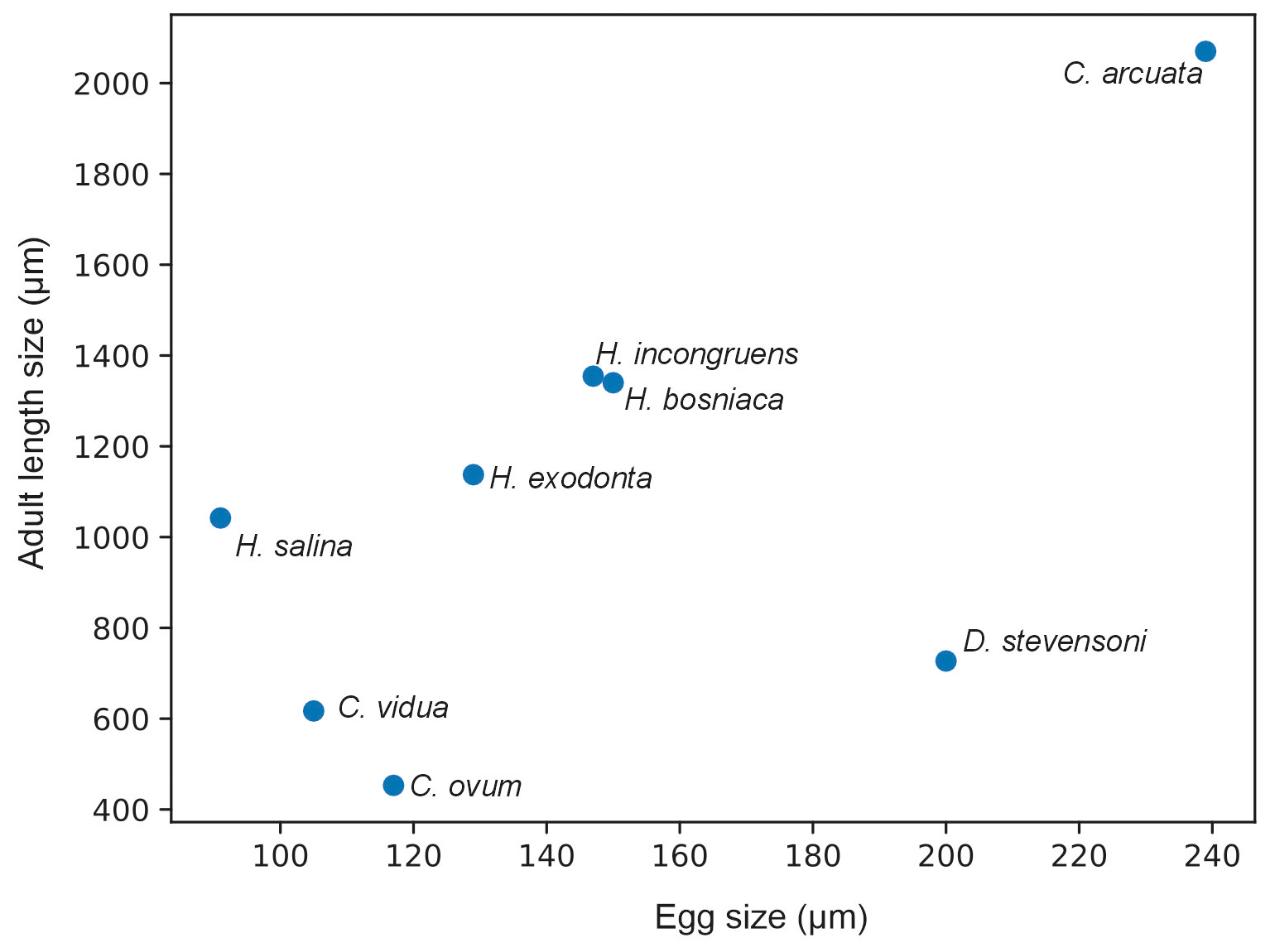
Figure 14Relationship between egg size (µm) and adult valve length (µm) in selected non-marine ostracod species based on data from this study and the literature. Adult size corresponds to the mean valve length (RV and LV when available). The plot shows a general positive tendency between egg and adult size, although the relationship is not strictly proportional across taxa and should be interpreted with caution.
This applies to both ecological and paleoecological studies. The use of different juvenile instars and adult stages in future paleoecological interpretations is crucial for integrative knowledge of ostracods, increasing their use as paleo-bioindicators. This approach allows for an appreciation of population structure, coupled with knowledge of relative abundances, facilitating comprehension of ostracod taphonomy (Whatley, 1988; Boomer et al., 2003). Consequently, discerning ontogeny assists in deciphering spatial and temporal alterations in sedimentation rates and transport magnitude, regulated by wind-induced hydrodynamics (Zhai et al., 2013; Zhai et al., 2015). Additionally, it is important to consider the fact that changes in population structure can be related to sediment type and aquatic ecosystem depth, factors that may influence ostracod distribution (Zhai et al., 2010).
Laboratory cultures of Heterocypris exodonta, H. incongruens, and H. salina made the documentation of complete ontogenetic sequences possible, including eight juvenile instars and the adult stage. The egg stage plays a key role in persistence under desiccation in ephemeral habitats. This resilience cannot be attributed solely to the eggshell but rather to the entire egg, which is adapted to enter dormancy. The eggshell, composed of an inner and an outer layer, likely contributes to protection against mechanical damage and environmental stress. Particular attention should be paid to the external egg surface as its morphology is not uniform across species and may represent a potentially informative taxonomic character in some taxa.
Ontogenetic changes in valve reticulation show a progressive reduction in ornamentation towards adulthood, highlighting the risk of misinterpretation when isolated valves are analyzed. The limited variation in growth factors further indicates that size ratios alone are not reliable diagnostic characters for closely related taxa. Importantly, laboratory cultures proved to be indispensable for capturing all developmental stages, which are rarely represented in full under natural conditions. By providing a complete ontogenetic framework, this study helps to distinguish ontogenetic variation from true interspecific differences, thereby improving taxonomic resolution and the interpretation of modern and fossil assemblages.
The morphometric measurements supporting this study are provided as a Supplement in Excel format associated with the article. Alternatively, the data are available in the doctoral thesis of Mauricio Bonilla Flores, published online by Technische Universität Braunschweig and accessible at https://doi.org/10.24355/dbbs.084-202409201141-0 (Bonilla Flores, 2025).
The supplement related to this article is available online at https://doi.org/10.5194/jm-45-429-2026-supplement.
Mauricio Bonilla-Flores: conceptualization, formal analysis, investigation, data curation, writing (original draft). Liseth Pérez: conceptualization, supervision, visualization, methodology, writing (review and editing). Peter Frenzel: methodology, supervision, writing (review and editing). Paula Echeverría-Galindo: conceptualization, methodology, investigation, resources, writing (review and editing). Junbo Wang: supervision, methodology, writing (review and editing). Antje Schwalb: conceptualization, funding acquisition, resources, writing (review and editing).
The contact author has declared that none of the authors has any competing interests.
Publisher's note: Copernicus Publications remains neutral with regard to jurisdictional claims made in the text, published maps, institutional affiliations, or any other geographical representation in this paper. The authors bear the ultimate responsibility for providing appropriate place names. Views expressed in the text are those of the authors and do not necessarily reflect the views of the publisher.
We thank all the colleagues from Nam Co Observation and Research Station for Multisphere (NAMORS), Institute of Tibetan Plateau Research, Chinese Academy of Sciences, for the support and assistance. We thank Simone Schulze, Faculty of Mechanical Engineering, TU Braunschweig, for taking the SEM images of the ostracods. We acknowledge the support of Nicole Börner, Sten Anslan, and Wengang Kang during the field sampling campaign. We thank Michael Kraft, Director of the Botanical Garden of TU Braunschweig, and Thorsten Marschall for allowing us to collect ostracods in the greenhouses and outdoor gardens. Thanks are given to Janine Melzig and Nora Kraatz for their assistance with sorting, counting, and measuring the ostracod valves. We are grateful to Francesc Mesquita-Joanes for providing the literature and to Dayou Zhai for their comments on growth ratios. Finally, we thank the anonymous reviewer and Renate Matzke-Karasz for their valuable comments on the paper.
This work was supported by the Deutsche Forschungsgemeinschaft (DFG) through the project “Geo-ecosystems in transition on the Tibetan Plateau” (TransTiP; grant no. 317513741/GRK2309).
This open-access publication was funded by Technische Universität Braunschweig.
This paper was edited by Moriaki Yasuhara and reviewed by Renate Matzke-Karasz and one anonymous referee.
Aguilar-Alberola, J. A. and Mesquita-Joanes, F.: Ontogeny of Heterocypris bosniaca (Ostracoda: Cyprididae): description of postembryonic instars and rediscovery of the neglected A-9 stage, J. Crustacean Biol., 33, 348–371, 2013.
Aguilar-Alberola, J. A. and Mesquita-Joanes, F.: Breaking the temperature–size rule: thermal effects on growth, development and fecundity of a crustacean from temporary waters, J. Therm. Biol., 42, 15–24, https://doi.org/10.1016/j.jtherbio.2014.02.016, 2014.
Aguilar-Alberola, J. A. and Mezquita, F.: On the presence of parthenogenetic populations of Heterocypris bosniaca Petkovski et al. 2000 (Crustacea: Ostracoda) in temporary rock pools of the eastern Iberian Peninsula, with insights into the ecology and life history of the species, Bull. Soc. Nat. Luxemb., 109, 149–158, 2008.
Akita, L. G., Frenzel, P., Wang, J., Börner, N., and Peng, P.: Spatial distribution and ecology of recent Ostracoda from Tangra Yumco and adjacent waters on the southern Tibetan Plateau: a key to palaeoenvironmental reconstruction, Limnologica, 59, 21–43, https://doi.org/10.1016/j.limno.2016.03.005, 2016.
Anderson, F. W.: The law of ostracod growth, Palaeontology, 7, 85–104, 1964.
Angell, R. W. and Hancock, J. W.: Response of eggs of Heterocypris incongruens (Ostracoda) to experimental stress, J. Crustac. Biol., 9, 381–386, https://doi.org/10.1163/193724089X00340, 1989.
Anslan, S., Azizi Rad, M., Buckel, J., Echeverria Galindo, P., Kai, J., Kang, W., Keys, L., Maurischat, P., Nieberding, F., Reinosch, E., Tang, H., Tran, T. V., Wang, Y., and Schwalb, A.: Reviews and syntheses: How do abiotic and biotic processes respond to climatic variations in the Nam Co catchment (Tibetan Plateau)?, Biogeosciences, 17, 1261–1279, https://doi.org/10.5194/bg-17-1261-2020, 2020.
Baird, W.: Arrangement of the British Entomostraca, with a list of species, particularly noticing those which have as yet been discovered within the bounds of the Club, Trans. Berwickshire Nat. Club., 2, 145–158, 1845.
Baltanás, A., Otero, M., Arqueros, L., Rossetti, G., and Rossi, V.: Ontogenetic changes in the carapace shape of the non-marine ostracod Eucypris virens (Jurine), Hydrobiologia, 419, 65–72, 2000.
Bellin, N., Groppi, M., and Rossi, V.: A model of egg bank dynamics in ephemeral ponds, Ecol. Model., 430, 109126, https://doi.org/10.1016/j.ecolmodel.2020.109126, 2020.
Bonilla Flores, M.: Integrative study of ostracods in high-altitude aquatic ecosystems, Nam Co, Tibetan Plateau: Taxonomic revisions, discovery of a new species, and ontogenetic features, Braunschweig, https://doi.org/10.24355/dbbs.084-202409201141-0, 2025.
Bonilla-Flores, M., Echeverría-Galindo, P., Frenzel, P., Pérez, L., Börner, N., Dulias, K., Wang, J., and Schwalb, A.: Taxonomy, ontogeny and ecology of Tonnacypris stewarti (Daday, 1908) comb. nov. (Ostracoda: Cyprididae) from Nam Co, Tibetan Plateau, Zootaxa, 5446, 331–365, https://doi.org/10.11646/zootaxa.5446.3.2, 2024.
Bonilla-Flores, M., Karanovic, I., Echeverría-Galindo, P., Frenzel, P., Pérez, L., Börner, N., Dulias, K., Wang, J., and Schwalb, A.: Heterocypris exodonta sp. nov. (Ostracoda: Cyprididae): morphological and molecular description of a high-altitude asexual microcrustacean from the Nam Co region, southern Tibetan Plateau, ZooKeys, 1264, 207–248, https://doi.org/10.3897/zookeys.1264.140174, 2025.
Boomer, I., Horne, D. J., and Slipper, I. J.: The use of ostracods in palaeoenvironmental studies, or what can you do with an ostracod shell?, Palaeontol. Soc. Pap., 9, 153–180, https://doi.org/10.1017/S1089332600002199, 2003.
Bouvier, C., Didon, E., Niedzielski, H. and Huizar, R.: Hidrología e hidrogeología en la cuenca de Chalco, descripción y perspectivas. ContactoS, 10, 19–29, 1993.
Brady, G. S.: A monograph of the recent British Ostracoda, Trans. Linn. Soc. Lond., 26, 353–495, https://doi.org/10.1080/00222936808695855, 1868.
Bronstein, Z. S.: Ostracoda presnykh vod. Fauna SSSR, Rakoobraznye, 2(1). Zoologicheskii Institut, Akademiya Nauk SSSR, Moscow. English translation: 1988. Fresh-water Ostracoda, Fauna of the USSR, Crustaceans, Amerind Publishing Co. Pvt. Ltd., New Delhi, xv + 470 pp., 1947.
Brooks, W. K.: Report on the Stomatopoda dredged by HMS Challenger during the years 1873–1876, Rep. Sci. Results Voy. HMS Challenger, Zoology 16, 1–116, https://doi.org/10.5962/bhl.title.9891, 1886.
Claus, C.: Beiträge zur Kenntnis der Süßwasser-Ostracoden: 1. Über den Körper- und Gliedmaßenbau der Cypriden nebst Bemerkungen über einzelne innere Organe derselben, Arbeiten aus dem Zoologischen Instituten der Universität Wien und der Zoologischen Station in Triest, 10, 147–216, 1892.
Danielopol, D. L., Baltanás, A., Namiotko, T., Geiger, W., Pichler, M., Reina, M., and Roidmayr, G.: Developmental trajectories in geographically separated populations of non-marine ostracods: morphometric applications for palaeoecological studies, Senckenberg. Lethaea, 88, 183–193, https://doi.org/10.1007/BF03043988, 2008.
De Deckker, P. and Martens, K.: Extraordinary morphological changes in valve morphology during the ontogeny of several species of the Australian ostracod genus Bennelongia (Crustacea, Ostracoda), Eur. J. Taxon., 36, 1–37, https://doi.org/10.5852/ejt.2013.36, 2013.
Delorme, D.: 20 Ostracoda. Ecology and classifications of North American Freshwater Invertebrates, 2nd Edn., https://doi.org/10.1016/B978-012690647-9/50021-1, 2011.
de Stasio, B. T.: Egg bank formation by aquatic invertebrates: a bridge across disciplinary boundaries, in: Diapause in aquatic invertebrates: theory and human use, edited by: Alekseev, V. R., de Stasio, B. T., and Gilbert, J. J., Springer, Dordrecht, 121–133, https://doi.org/10.1007/978-1-4020-5680-2_7, 2007.
Díaz, A. R. and Lopretto, E. C.: Postembryonic development of the non-marine ostracod Chlamydotheca arcuata (Sars, 1901) reared in the laboratory, Turk. J. Zool., 41, 209–226, https://doi.org/10.3906/zoo-1512-13, 2017.
Dumont, H. J., Nandini, S., and Sarma, S. S. S.: Cyst ornamentation in aquatic invertebrates: a defence against egg predation, Hydrobiologia, 486, 161–167, https://doi.org/10.1023/A:1021346601235, 2002.
Fontana, S. L. and Ballent, S.: A new giant cypridid ostracod (Crustacea) from southern Buenos Aires Province, Argentina, Hydrobiologia, 533, 187–197, https://doi.org/10.1007/s10750-004-2415-8, 2005.
Fryer, G.: The horse-trough ostracod Heterocypris incongruens, Naturalist, 122, 121–135, 1997.
Hairston Jr., N. G. and Fox, J. A.: Egg banks, in: Encyclopedia of Inland Waters, edited by: Likens, G. E., Academic Press, Oxford, 659–666, https://doi.org/10.1016/B978-012370626-3.00152-6, 2009.
Hartmann, G.: Ostracoda, in: Klassen und Ordnungen des Tierreichs, Vol. 5: Arthropoda, Teil Crustacea, edited by: Gruner, H.-E., Akademische Verlagsgesellschaft Geest & Portig K.-G., Leipzig, 409–568, 1968.
Herman, P. M. J. and Heip, C.: Growth and respiration of Cyprideis torosa (Jones, 1850) (Crustacea, Ostracoda), Oecologia, 54, 300–303, https://doi.org/10.1007/BF00379996, 1982.
Holmes, J. A.: Ostracoda, in: Tracking environmental change using lake sediments, Vol. 4, edited by: Smol, J. P., Birks, H. J. B., and Last, W. M., Kluwer Academic Publishers, Dordrecht, 125–151, https://doi.org/10.1007/0-306-47671-1_7, 2001.
Horne, D. J. and Martens, K.: An assessment of the importance of resting eggs for the evolutionary success of Mesozoic non-marine cypridoidean Ostracoda (Crustacea), Arch. Hydrobiol. Spec. Issues Advanc. Limnol, 52, 549–561, 1998.
Horne, D. J., Smith, R. J., Whittaker, J. E., and Murray, J. W.: The first British record and a new species of the superfamily Terrestricytheroidea (Crustacea, Ostracoda): morphology, ontogeny, lifestyle and phylogeny, Zool. J. Linn. Soc., 142, 253–288, https://doi.org/10.1111/j.1096-3642.2004.00134.x, 2004.
Karanovic, I. and Lee, W.: A review of candonid ostracods (Crustacea: Ostracoda: Podocopida) from East Asia, with descriptions of five new species from South Korea, Zootaxa, 3368, 7–49, 2012.
Kesling, R. V.: The morphology of ostracod molt stages, Ill. Biol. Monogr., 21, 1–126, 1951.
Kesling, R. V.: Doubling in size of ostracod carapaces in each molt stage, J. Paleontol., 26, 772–780, 1952.
Kesling, R. V. and Crafts, F. C.: Ontogenetic increase in Archimedean weight of the ostracod Chlamydotheca unispinosa (Baird), Am. Midl. Nat., 68, 149–153, https://doi.org/10.2307/2422641, 1962.
Kilikowska, A., Schön, I., Wysocka, A., Pieri, V., Kaczmarczyk-Ziemba, A., Prais, K., Martens, M., and Namiotko, T.: Multiple genetic species in a halophilic non-marine ostracod (Crustacea), Hydrobiologia, 851, 2447–2467, https://doi.org/10.1007/s10750-023-05469-z, 2024.
Kong, Q., Karanovic, I. and Yu, N.: Phylogeny of the genus Chrissia (Ostracoda: Cyprididae) with description of a new species from China, J. Crustac. Biol., 34, 782–794, https://doi.org/10.1163/1937240X-00002276, 2014.
Kubanç, N., Özuluğ, O., and Kubanç, C.: Ontogeny of appendages of Heterocypris salina (Brady, 1868) (Crustacea, Ostracoda), Hydrobiologia, 585, 255–272, https://doi.org/10.1007/s10750-007-0642-5, 2007.
Lajblová, K., Kraft, P., and Meidla, T.: Ontogeny of the ostracod Conchoprimitia osekensis (Přibyl, 1979) from the Darriwilian of the Prague Basin (Czech Republic), Est. J. Earth Sci., 63, 144–155, https://doi.org/10.3176/earth.2014.13, 2014.
Latreille, P. A.: Histoire naturelle, générale et particulière des Crustacés et des Insectes. Ouvrage faissant suite à l'histoire naturelle générale et particulière, composée par Leclerc de Buffon, et rédigée par C.S. Sonnini, membre de plusieurs sociétés savantes, Dufart, Paris, 467 pp., 1802.
Liberto, R., César, I. I., and Mesquita-Joanes, F.: Postembryonic growth in two species of freshwater Ostracoda (Crustacea) shows a size–age sigmoid model fit and temperature effects on development time, but no clear temperature–size rule (TSR) pattern, Limnology, 15, 57–67, https://doi.org/10.1007/s10201-013-0413-0, 2014.
Mao, X., Liu, X., Li, J., Feng, S., Jiang, G., and Liu, L.: Population age structure of ostracods in lake sediment and its implication for within-lake transport of microfossils, Ecol. Indic., 131, 108182, https://doi.org/10.1016/j.ecolind.2021.108182, 2021.
Marín, J. A.: Estudio del desarrollo de los ostrácodos Eucypris aragonica y Heterocypris salina en cultivo de barro, Limnetica, 1, 345–354, 1984.
Martens, K.: Effects of temperature and salinity on postembryonic growth in Mytilocypris henricae (Chapman) (Crustacea, Ostracoda), J. Crustac. Biol., 5, 258–272, 1985.
Matzke-Karasz, R., Smith, R. J., and Heß, M.: Removal of extracellular coat from giant sperm in female receptacle induces sperm motility in Mytilocypris mytiloides (Cyprididae, Ostracoda, Crustacea), Cell Tissue Res., 368, 171–186, https://doi.org/10.1007/s00441-016-2507-6, 2017.
McCormack, J., Viehberg, F., Akdemir, D., Immenhauser, A., and Kwiecien, O.: Ostracods as ecological and isotopic indicators of lake water salinity changes: the Lake Van example, Biogeosciences, 16, 2095–2114, https://doi.org/10.5194/bg-16-2095-2019, 2019.
Meisch, C.: Freshwater Ostracoda of Western and Central Europe, Süßwasserfauna von Mitteleuropa 8/3, Spektrum Akademischer Verlag, Heidelberg, 522 pp., ISBN 3-8274-1001-0, 2000.
Meisch, C., Smith, R. J., and Martens, K.: An updated subjective global checklist of the extant non-marine Ostracoda (Crustacea), Eur. J. Taxon., 974, 1–144, https://doi.org/10.5852/ejt.2024.974.2767, 2024.
Mesquita-Joanes, F., Aguilar-Alberola, J. A., Palero, F., and Rueda, J.: A new species of Cypris (Crustacea: Ostracoda) from the Iberian Peninsula and the Balearic Islands, with comments on the first ostracod named using the Linnean system, Zootaxa, 4759, 113–131, https://doi.org/10.11646/zootaxa.4759.1.8, 2020.
Mezquita, F., Griffiths, H. I., Sanz, S., Soria, J. M., and Piñón, A.: Ecology and distribution of ostracods associated with flowing waters in the eastern Iberian Peninsula, J. Crustac. Biol., 19, 344–354, 1999.
Mezquita, F., Olmos, V., and Oltra, R.: Population ecology of Cyprideis torosa (Jones, 1850) in a hypersaline environment of the Western Mediterranean (Santa Pola, Alacant) (Crustacea: Ostracoda), Ophelia, 53, 119–130, https://doi.org/10.1080/00785236.2000.10409442, 2000.
Mezquita, F., Boronat, M. D., and Miracle, M. R.: The life history of Cyclocypris ovum (Ostracoda) in a permanent karstic lake, Arch. Hydrobiol., 155, 687–704, 2002.
Müller-Calé, C.: Über die Entwicklung von Cypris incongruens, Zool. Jahrb., Abt. Anat. Ont. Tiere, 36, 113–170, 1913.
Oakley, T. H., Wolfe, J. M., Lindgren, A. R., and Zaharoff, A. K.: Phylotranscriptomics to bring the understudied into the fold: monophyletic Ostracoda, fossil placement and pancrustacean phylogeny, Mol. Biol. Evol., 30, 215–233, https://doi.org/10.1093/molbev/mss216, 2012.
Özuluğ, O. and Suludere, Z.: Morphology of dry-resistant eggs in parthenogenetic Heterocypris incongruens (Ramdohr, 1808) (Crustacea, Ostracoda), Acta Biol. Hung., 63, 333–341, https://doi.org/10.1556/ABiol.63.2012.3.3, 2012.
Pereira, J. S., Schneider, D. I. D., da Rocha, C. E. F., and Pinto, R. L.: Carapace ontogeny of the bromeliad-dwelling ostracod Elpidium bromeliarum Müller, 1880 (Crustacea, Ostracoda), J. Nat. Hist., 51, 1–12, https://doi.org/10.1080/00222933.2017.1360529, 2017.
Petkowski, T., Scharf, B., and Keyser, D.: New and little known ostracods of the genus Heterocypris (Crustacea, Ostracoda) from the Balkan Peninsula, Limnologica, 30, 45–57, https://doi.org/10.1016/S0075-9511(00)80042-0, 2000.
Ramdohr, A. F.: Über die Gattung Cypris Müller und drei zu derselben gehörige neue Arten, Mag. Ges. Naturf. Fr. Berlin Neusten Entdeck Ges. Naturkd., 2, 85–93, 1808.
Ranta, E.: Population biology of Darwinula stevensoni (Crustacea, Ostracoda) in an oligotrophic lake, Ann. Zool. Fenn., 16, 28–35, 1979.
R Core Team: R: a language and environment for statistical computing, R Foundation for Statistical Computing, Vienna, https://www.R-project.org/ (last access: 15 January 2026), 2022.
Retrum, J. B. and Kaesler, R. L.: Early Permian Carbonitidae (Ostracoda): ontogeny, affinity, environment and systematics, J. Micropalaeontol., 24, 179–190, https://doi.org/10.1144/jm.24.2.179, 2005.
Roessler, E. W.: Estudios taxonómicos, ontogenéticos, ecológicos y etológicos sobre los ostrácodos de agua dulce en Colombia – II. Contribución al conocimiento del desarrollo embrionario tardío y de los procesos de la eclosión del huevo de Heterocypris bogotensis Roessler (Ostracoda, Podocopa, Cyprididae), Caldasia, 13, 453–466, http://www.jstor.org/stable/23641185 (last access: 25 April 2026), 1982.
Roessler, E. W.: Estudios taxonómicos, ontogenéticos, ecológicos y etológicos sobre los ostrácodos de agua dulce en Colombia – IV. Desarrollo postembrionario de Heterocypris bogotensis Roessler (Ostracoda, Podocopa, Cyprididae), Caldasia, 13, 755–776, https://www.jstor.org/stable/23641092 (last access: 25 April 2026), 1983.
Rosa, J., Campos, R., Martens, K., and Higuti, J.: Spatial variation of ostracod (Crustacea, Ostracoda) egg banks in temporary lakes of a tropical floodplain, Mar. Freshw. Res., 72, 26–34, https://doi.org/10.1071/MF19081, 2020.
Rosa, J., Petsch, D. K., Martens, K., and Higuti, J.: Species' traits and taxonomic distance can predict the hatching phenology of ostracod (Crustacea) resting eggs from tropical floodplain lakes, Int. Rev. Hydrobiol., 106, 226–238, https://doi.org/10.1002/iroh.202102105, 2021.
Rosa, J., Martens, K., and Higuti, J.: Dried aquatic macrophytes are floating egg banks and potential dispersal vectors of ostracods (Crustacea) from pleuston communities, Hydrobiologia, 850, 1319–1329, https://doi.org/10.1007/s10750-022-04818-8, 2023.
Rossi, V. and Menozzi, P.: Inbreeding and outbreeding depression in geographical parthenogens Heterocypris incongruens and Eucypris virens (Crustacea, Ostracoda), Ital. J. Zool., 79, 559–567, https://doi.org/10.1080/11250003.2012.718375, 2012.
Rossi, V., Gandolfi, A., and Menozzi, P.: Egg diapause and clonal structure in parthenogenetic populations of Heterocypris incongruens (Ostracoda), Hydrobiologia, 320, 45–54, 1996.
Rossi, V., Tondelli, B., Gandolfi, A., Bellavere, C., McKenzie, K. G., and Menozzi, P.: Heterocypris (Crustacea, Ostracoda) from the Isole Pelagie (Sicily, Italy): population genetics, Ital. J. Zool., 70, 101–108, https://doi.org/10.1080/11250000309356502, 2003.
Rossi, V., Benassi, G., Leonardi, S., Piotti, A., and Menozzi, P.: Clonal diversity of Heterocypris incongruens (Crustacea: Ostracoda) in Northern Italian ricefields, Arch. Hydrobiol., 166, 225–240, https://doi.org/10.1127/0003-9136/2006/0166-0225, 2006.
Rossi, V., Gandolfi, A., Baraldi, F., Bellavere, C., and Menozzi, P.: Phylogenetic relationships of coexisting Heterocypris (Crustacea, Ostracoda) lineages with different reproductive modes from Lampedusa Island (Italy), Mol. Phylogenet. Evol., 44, 1273–1283, https://doi.org/10.1016/j.ympev.2007.04.013, 2007.
Rossi, V., Albini, D., Benassi, G., and Menozzi, P.: To rest in hydration: hatching phenology of resting eggs of Heterocypris incongruens (Crustacea, Ostracoda), Fundam. Appl. Limnol., 181, 49–58, https://doi.org/10.1127/1863-9135/2012/0251, 2012.
Rossi, V., Zattori, M., Perotti, A., and Menozzi, P.: What does egg size tell us about hatching phenology in Heterocypris incongruens?, Fundam. Appl. Limnol., 182, 253–270, 2013.
Rossi, V., Albini, D., Pellegri, V., and Menozzi, P.: Early and late maternal effects on hatching phenology of Heterocypris incongruens (Crustacea: Ostracoda), J. Exp. Zool. A, 323, 382–391, https://doi.org/10.1002/jez.1931, 2015.
Sars, G. O.: Oversigt af Norges marine ostracoder. [Overview of Norway's marine ostracods], Forhandlinger i Videnskabs-Selskabet i Christiania, 1865, 1–130, 1866.
Sars, G. O.: Contribution to the knowledge of the freshwater Entomostraca of South America as shown by artificial hatching from dried material. Arch. Math. Naturvidensk, 24, 3–52, 1901.
Schmit, O., Rossetti, G., Vandekerkhove, J., and Mezquita, F.: Food selection in Eucypris virens (Crustacea: Ostracoda) under experimental conditions, Hydrobiologia, 585, 135–140, https://doi.org/10.1007/s10750-007-0634-5, 2007.
Schreiber, E.: Beiträge zur Kenntnis der Morphologie, Entwicklung und Lebensweise der Süsswasser-Ostracoden, Zool. Jahrb., 43, 485–538, 1922.
Shaver, R. H.: Ontogeny and sexual dimorphism in Cytherella bullata, J. Paleontol., 27, 471–480, 1953.
Smith, A. J., Horne, D. J., Martens, K., and Schön, I.: Class Ostracoda, in: Thorp and Covich's Freshwater Invertebrates, 4th Edn., edited by: Thorp, J. H. and Rogers, D. C., Academic Press, 757–780, https://doi.org/10.1016/B978-0-12-385026-3.00030-9, 2015.
Smith, R. J. and Kamiya, T.: Ontogeny of Neonesidea oligodentata (Ostracoda), Hydrobiologia, 489, 245–275, https://doi.org/10.1023/A:1023221303564, 2002.
Smith, R. J. and Kamiya, T.: Copulatory behaviour and sexual morphology of three Fabaeformiscandona Krstić, 1972 (Candoninae, Ostracoda, Crustacea) species from Japan, including descriptions of two new species, Hydrobiologia, 585, 225–248, https://doi.org/10.1007/s10750-007-0640-7, 2007.
Smith, R. J. and Martens, K.: The ontogeny of the cypridid ostracod Eucypris virens (Jurine, 1820) (Crustacea, Ostracoda), Hydrobiologia, 419, 31–63, https://doi.org/10.1023/A:1003985908460, 2000.
Song, J. J., Huang, B., and Qie, W. K.: Allometry in Late Devonian podocopid ostracods and its implications for ostracod ontogeny, Papers Palaeontol., e1480, https://doi.org/10.1002/spp2.1480, 2023.
Tinn, O. and Meidla, T.: Ontogeny and thanatocoenoses of early Middle Ordovician palaeocope ostracods, J. Paleontol., 77, 64–72, https://doi.org/10.1666/0022-3360(2003)077<0064:OATOEM>2.0.CO;2, 2003.
Vandekerkhove, J., Namiotko, T., Hallmann, E., and Martens, K.: Predation by macroinvertebrates on Heterocypris incongruens (Ostracoda) in temporary ponds: impacts and responses, Fundam. Appl. Limnol., 181, 39–47, https://doi.org/10.1127/1863-9135/2012/0310, 2012.
Van Doninck, K., Schön, I., Martens, K., and Goddeeris, B.: The life cycle of the asexual ostracod Darwinula stevensoni (Brady and Robertson, 1870), Hydrobiologia, 500, 331–340, https://doi.org/10.1023/A:1024656920904, 2003.
Vences, M., Anslan, S., Sabino-Pinto, J., Bonilla-Flores, M., Echeverría-Galindo, P., John, U., Nass, B., Pérez, L., Preick, M., Zhu, L., and Schwalb, A.: Dataset from RNA-seq analysis of differential gene expression among developmental stages of two non-marine ostracods, Data Brief, 53, 110070, https://doi.org/10.1016/j.dib.2024.110070, 2024.
Wang, C., Wang, H., Kuang, X., and Guo, G.: Life stages and morphological variations of Limnocythere inopinata (Crustacea, Ostracoda) from Lake Jiang-Co (northern Tibet), ZooKeys, 1011, 25–40, https://doi.org/10.3897/zookeys.1011.56065, 2021.
Wang, Q., Zhai, D., Peng, A., Schmitz, O., Jiang, P., Fang, X., Wang, M., and Frenzel, P.: Adapted to the dry: ostracod communities in agricultural landscapes of eastern Yunnan, China, Aquat. Ecol., 59, 541–559, https://doi.org/10.1007/s10452-025-10179-w, 2025.
Watabe, K. and Kaesler, R. L.: Ontogeny of a new species of Paraparchites (Ostracoda) from the Lower Permian of Kansas, J. Paleontol., 78, 603–611, https://doi.org/10.1666/0022-3360(2004)078<0603:OOANSO>2.0.CO;2, 2004.
Weygoldt, P.: Embryologische Untersuchungen an Ostrakoden: die Entwicklung von Cyprideis litoralis (G. S. Brady) (Ostracoda, Podocopa, Cytheridae), Zool. Jahrb. Abt. Anat. Ontog. Tiere, 78, 367–426, 1960.
Whatley, R. C.: Population structure of ostracods: some general principles for the recognition of palaeoenvironments, in: Ostracoda in the Earth Sciences, edited by: De Deckker, P., Colin, J. P., and Peypouquet, J. P., Elsevier, Amsterdam, 245–256, , 1988.
Wohlgemuth, R.: Beobachtungen und Untersuchungen über die Biologie der Süsswasserostracoden: ihr Vorkommen in Sachsen und Böhmen, ihre Lebensweise und ihre Fortpflanzung, J. Klinkhardt, Leipzig, 1914.
Woltereck, R.: Zur Bildung und Entwicklung des Ostrakoden-Eies: kerngeschichtliche und biologische Studien an parthenogenetischen Cypriden, Z. Wiss. Zool., 64, 595–623, 1898.
Yavuzatmaca, M. and Külköylüoğlu, O.: Sexual populations of Heterocypris incongruens (Ramdohr, 1808) from Turkey, Acta Aquat. Turc., 15, 139–150, https://doi.org/10.22392/actaquatr.577460, 2019.
Yoo, H., Cohuo, S., Macario-González, L., and Karanovic, I.: A new freshwater ostracod genus from the northern Neotropical region and its phylogenetic position within Cyprididae, Zool. Anz., 266, 196–215, https://doi.org/10.1016/j.jcz.2016.09.003, 2017.
Yousef, E. A. and Alahmadi, B. A.: Fecundity, egg volume, and reproductive output of Xestoleberis rotunda (Podocopida: Ostracoda) from the Red Sea, Egypt, Egypt. J. Aquat. Res., 51, 189–197, https://doi.org/10.1016/j.ejar.2025.01.007, 2025.
Yousef, E. A., Seleem, A. A., and Alansari, R. M.: Ontogeny changes in carapace and appendages of ostracod Cypris pubera Müller, 1776, Egypt. J. Aquat. Res., 50, 472–489, https://doi.org/10.1016/j.ejar.2024.09.005, 2024.
Zhai, D., Xiao, J., Zhou, L., Wen, R., Chang, Z., and Pang, Q.: Similar distribution patterns of different phenotypes of Limnocythere inopinata in a brackish-water lake, Hydrobiologia, 651, 185–197, https://doi.org/10.1007/s10750-010-0295-7, 2010.
Zhai, D., Xiao, J., Fan, J., Zhou, L., Wen, R., and Pang, Q.: Spatial heterogeneity of the population age structure of Limnocythere inopinata in Hulun Lake, Hydrobiologia, 716, 29–46, https://doi.org/10.1007/s10750-013-1541-6, 2013.
Zhai, D., Xiao, J., Fan, J., Wen, R., and Pang, Q.: Differential transport and preservation of instars of Limnocythere inopinata, Hydrobiologia, 747, 1–18, https://doi.org/10.1007/s10750-014-2118-8, 2015.
Zhu, L., Wu, Y., Wang, J., Lin, X., Ju, J., Xie, M., Li, M., Mäusbacher, R., Schwalb, A., and Daut, G.: Environmental changes since 8.4 ka reflected in lacustrine sediments from Nam Co, central Tibetan Plateau, Holocene, 18, 831–839, 2008.